Article Text

Abstract
Background: The mucosal immune system undergoes extensive changes in early childhood in response to environmental stimuli. Dendritic cells (DC) play a major role in the development of the immune system. However, few data exist on the influence of continuous environmental stimulation on the distribution and phenotype of human airway DC.
Methods: Human tissue samples are mostly paraffin embedded which limits the use of several antibodies, and respiratory tissue for cryopreservation is difficult to obtain. Human frozen post mortem tracheal tissue was therefore used for this study. Only samples with epithelial adherence to the basement membrane were included (n = 34). Immunohistochemical staining and sequential overlay immunofluorescence were performed with DC-SIGN and a panel of leucocyte markers co-expressed by DC.
Results: DC detected in the human tracheal mucosa using DC-SIGN correlated with the expression of HLA-DR, co-stimulatory and adhesion molecules. Higher cell densities were found at the ventral tracheal site of patients older than 1 year than in infants in the first year of life.
Conclusion: The increasing population of mucosal DC with age could reflect immunological maturation.
- DC, dendritic cells
- SIDS, sudden infant death syndrome
- dendritic cells
- respiratory mucosa
- age
- acquired immunity
Statistics from Altmetric.com
Dendritic cells (DC) have an important role in the mucosal immune system of the respiratory tract, being involved in antigen uptake, processing, and presentation of antigenic fragments to T cells.1–3 Despite an enormous increase in knowledge about human DC derived from peripheral blood, the characterisation of these cells in human lymphoid and non-lymphoid tissues is still limited.4,5,6,7,8,9,10,11 DC have been characterised in mouse models in non-lymphoid organs such as the skin, gut and lung.12–14 The major relevance of DC in the uptake of particles in the bronchoalveolar space and the transport to the draining lymph node have recently been described, stressing the significant role of DC in the lung and their traffic to the lymph node.15 DC are important in the pathophysiology of asthma and might help to explain the “hygiene hypothesis”, as epidemiological observations suggest a vital role in the development of allergy and/or tolerance during early childhood.16,17 However, there are few human data on the presence and development of mucosal DC during this early period of life.18,19
A recent study suggested that adjuvant co-stimulation triggers oral tolerance mechanisms in neonatal mice.20 The development of neonatal immunity in respect to allergic diseases in childhood and in adults is of clinical relevance. DC are necessary for the initiation and modulation of T cell responses and are involved in immunoregulation early in life.21,22 An increase in DC after the first year of life has been observed in human tracheas.17 Animal studies have shown a distinct topographical distribution of DC between different layers of the tracheobronchial mucosa, although there are differences between species.23,24 The few data that exist on DC in the mucosa of the human respiratory tract are mainly derived from studies on tissues after surgery or biopsy specimens, generally from adult patients.25–28 Recent investigations on isolated lung DC using new antibodies (BDCA1, 2, 3) in flow cytometry have demonstrated three subsets of human DC—myeloid DC types 1 and 2 and plasmacytoid DC.29
In a previous study with paraffin embedded tissues, the presence of DC in the human trachea was examined using HLA-DR labelling and morphology because of a lack of antibodies staining paraffin sections.18 In a recent study DC were characterised, in addition to other leucocytes, in situ in frozen human post mortem tissue and resection material. In this investigation no differences in the cell numbers were found between post mortem and surgical samples.30 Vermaelen and Pauwels31 recently reviewed the current knowledge and unanswered questions on DC populations and their possible functions in the lung.
In previous studies, DC have been detected in cell suspensions of human bronchoalveolar lavage fluid and in minced human lung tissue. The morphological data on DC in human lung tissue are still limited. A study was therefore undertaken to examine the influence of age on the phenotype and predominant localisation of DC in the human tracheal mucosa in situ. A topographical and quantitative analysis was performed in human tissue samples using a set of recently available antibodies including DC-SIGN as a specific marker for DC.32
METHODS
Tissue collection
Tracheal tissue was obtained from the departments of Pathology and Legal Medicine of the Medical School of Hannover. Only samples with epithelial adherence to the basement membrane as assessed by haematoxylin staining (n = 26) were selected. Fresh resection material was provided by the Heidehaus Hospital, Hannover (n = 8) after partial transversal trachectomy. Details of the age, sex, and cause of death or surgical intervention are shown in tables 1 and 2.
Specification of included tissues: surgical specimens
Specification of included tissues: pathological and legal medicine specimens
The samples were taken from the lower part of the trachea near the carina (fig 1), immediately snap frozen in liquid nitrogen, and stored at −80°C. In addition, in some cases the upper part of the trachea was excised near the cricoid (n = 6). All tissues were free of tumours. Collection of the tissues was approved by the ethical committee of the Medical School of Hannover.
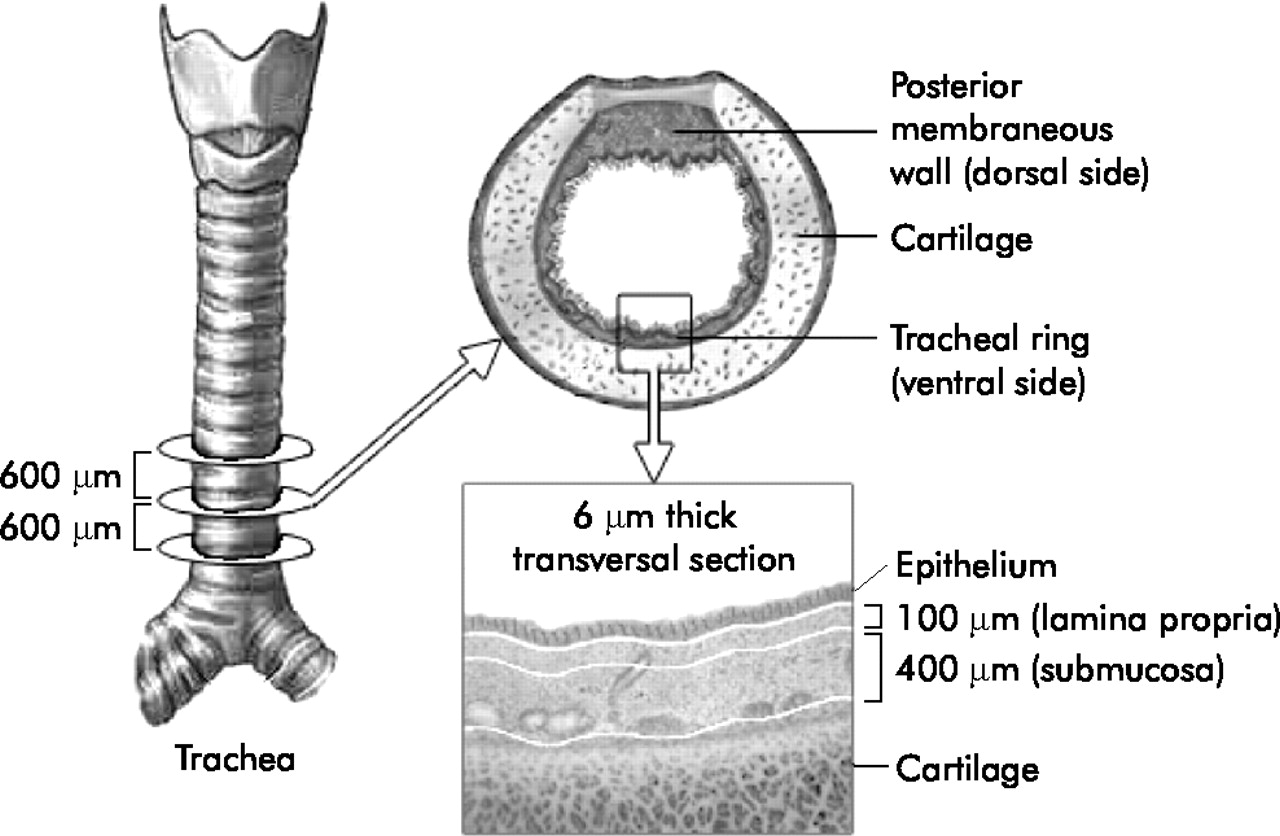
Transverse tracheal sections of 6 μm taken at intervals of 600 μm cranially were stained for a panel of markers. Analysis was performed on the ventral part of the trachea and only in the cartilage region to exclude possible variation due to location. The total mucosa is the combination of the lamina propria and submucosa. The epithelium, total mucosa, lamina propria and the submucosa were analysed separately.
Immunohistology
Serial cryosections of 6 μm (cryostat Leica CM3050S) on poly-L-lysin coated slides were stained for a large panel of immunological relevant DC markers (antibody/clone/dilution CD209-DC-SIGN/120507/1:20, R&D Systems Gmbh, Wiesbaden, Germany, CD11a/MHM24/1:100, CD86/BU63/1:20, HLA-DR/C3,43/1:10.000, Dako A/S, Glostrup, Denmark; CD11c/HL3/1:100, CD54/HA58/1:2500, BD Pharmingen, San Diego, CA, USA; CD40/LOB7,6/1:100, CD80/DAL-1/1:20, Serotec Ltd, Oxford, UK). Isotype controls (A57H, Dako A/S, Glostrup, Denmark) were included. Immunohistochemical staining was performed on acetone fixed sections using the APAAP method (secondary and detection antibody: Z0259 and D0651, Dako A/S) with FastBlue as substrate and counterstained with haematoxylin (Mayers’ haematoxylin, Merck KG, Darmstadt, Germany). Unfortunately, several other antibodies which are used in cytological applications for human DC do not work in histological preparations in general or in the post mortem material used in the present study.
For immunofluorescence staining, the same protocol and antibodies were used but after direct conjugation with Zenon Labeling Kits (Alexa Fluor 488 and 568, Molecular Probes, Leiden, The Netherlands) followed by a signal amplification step when necessary (Alexa Fluor 488 Signal-Amplification Kit for Fluorescein and Oregon Green Dye conjugated probes, Molecular Probes). Nuclei were stained with TO-PRO-3 (Molecular Probes). Owing to technical problems, not all antibodies could be applied to the sections of all patients. The total number of staining data for some antibodies is therefore less than the number of all 40 cases.
Analysis and statistics
Analysis of the immunohistochemically stained slides was performed using a light microsope (Axiophot, Zeiss, Oberkochen, Germany) with a grid at a 20× magnification. Pictures were recorded digitally (Olympus DP-Soft, version 3.2) with a fixed gamma correction and brightness. The immunofluorescent stained slides were also analysed at 20× magnification by means of confocal microscopy (LSM510meta, Leica, Germany). Cell densities (cells/mm2) were evaluated as previously described.29 The lamina propria (100 μm from the basement membrane) and submucosa (100–500 μm from the basement membrane) were analysed separately. Three sections of every marker each 600 μm apart were studied over a length of at least 10 mm basement membrane (fig 1). Areas with glands (intercartilage region) were excluded from these cell counts. Separate cell counts were performed for the posterior membranous wall (dorsal site) and the tracheal ring (ventral site) of the tracheas of infants (n = 6) and adults (n = 28), and also for the lamina propria and submucosa in single stained serially cut tissue slides for each marker.
The Mann-Whitney U test was used for comparison of non-parametric values and correlations were calculated using StatView Version 5.0 (SAS Institute Inc, Cary, NC, USA). All values are given as mean (SE). A p value of <0.05 was considered statistically significant.
RESULTS
Immunohistochemistry, immunofluorescence, and correlation of marker expression
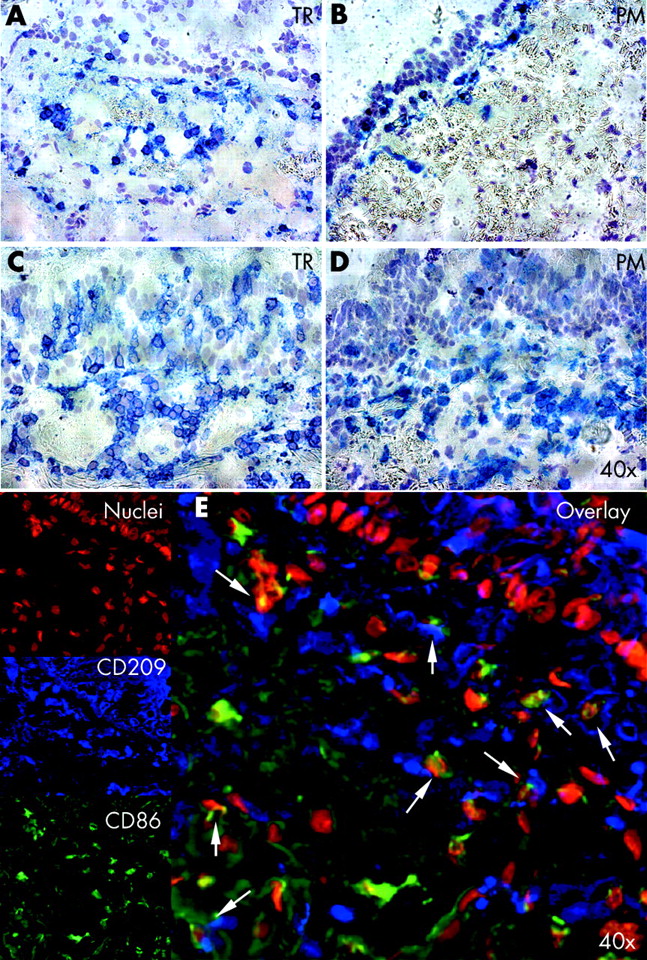
No significant differences in the number and subsets of DC were found in six cases where the upper and lower parts of the same trachea were examined, so only the lower part was studied in more detail in subsequent samples. The highest leucocyte and DC densities were found in the lamina propria just beneath the basement membrane (fig 2A–D). With the exception of CD80, all other investigated leucocyte surface molecules correlated with the expression of DC-SIGN (fig 3). Sequential immunofluorescence overlays showed that DC-SIGN positive cells expressed co-stimulatory and adhesion molecules (fig 2E). Although no correlation was found between CD80 and DC-SIGN, co-expression with DC-SIGN was found in the overlays. For CD54, focal staining was usually observed in the respiratory epithelium and the glands of the tracheal ring but not in the posterior membranous wall.

Immunohistochemistry and sequential overlay immunofluorescence. Comparison of cell densities for CD11a of infants (A, B: patient 26) and adults (C, D: patient 24) for the ventral trachea (TR) and dorsal trachea (posterior membranous wall, PM) at 40× magnification. (E) Sequential overlay immunofluorescent staining for CD209 (blue), CD86 (green), and nuclei (red) in patient 24 (magnification 20×).

(A–G) Correlations between CD209 and other dendritic cells (HLA-DR, CD11c) and leucocyte surface molecules (CD40, CD54, CD80, CD86) in the lamina propria. All axes represent cell densities in cell/mm2. Only the tissues in which all markers were analysed are included (six infants and 19 adults). All correlations were significant (p<0.05) except the correlation with CD80.
Age related differences
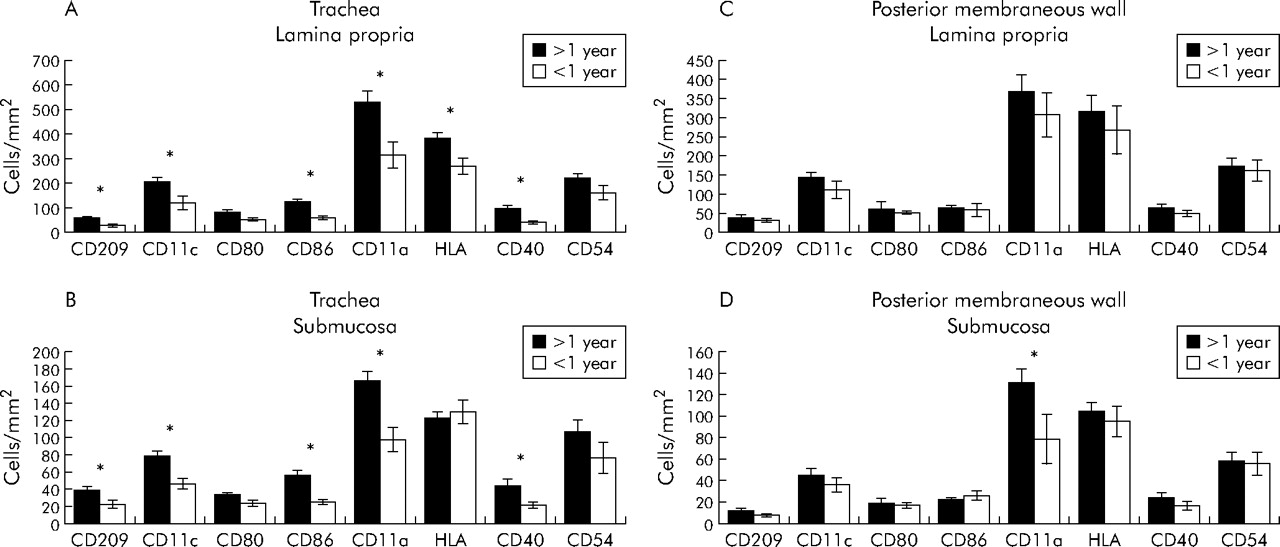
Age dependent differences were found in the distribution and abundance of cells in the tracheal ring (fig 4A, B). In the lamina propria, infants under the age of 1 year had lower cell densities for DC-SIGN, HLA-DR, CD11a/c, CD86, and CD40 than older patients. Except for HLA-DR, the results were similar in the submucosa. Interestingly, almost no differences were found between the two mucosal layers of the posterior membranous wall at the dorsal site (fig 4C, D). Only the cell density of CD11a expressing cells in the submucosa was higher in patients older than 1 year. No differences were found between prenatal deaths and infants diagnosed post mortem with sudden infant death.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cell densities for the lamina propria and submucosa in adults and infants in (A, B) the ventral part of the trachea (6 infants, 28 adults) and (C, D) the dorsal part of the trachea (5 infants, 14 adults). *p<0.05.
Comparison of distinct topographical sites of the trachea
For the comparison of site specific cell distribution, only cases with the tracheal ring (ventral) and the posterior membranous wall (dorsal) were included (n = 19). In patients older than 1 year of age, significantly (p<0.05) higher densities were found in the whole mucosa (lamina propria and submucosa) of the tracheal ring than in the posterior membranous wall for all markers (mean (SE): DC-SIGN 43 (3)/24 (3); CD11c 107 (8)/67 (6); CD80 45 (4)/29 (7); CD86 71 (6)/31 (3); CD11a 250 (16)/156 (16); CD40 56 (9)/32 (5); CD54 132 (14)/85 (11) cells/mm2). The numbers of HLA-DR did not differ significantly (182 (9)/154 (14), p = 0.052). This was also true for DC in the lamina propria and in the submucosa, but without reaching significance at the single layer level (fig 4). These differences were not seen in infants under the age of 1 year. For CD54, focal staining was usually observed in the respiratory epithelium and the glands of the tracheal ring but not in the posterior membranous wall.
DISCUSSION
Studies to date on human DC have used nasal, laryngeal, bronchial, and peripheral lung tissue but not tissue derived from the trachea.25–28,31,33,34 This study presents data on a large group of cryopreserved post mortem collected tracheal tissue and tissue collected from patients during surgery. Cryopreserved tissue offers the opportunity to use many monoclonal antibodies as markers. This was important since paraffin embedded tissue is not usable for most DC antibodies. The use of specific antibodies was necessary because the characteristic morphology of the DC is often not present in post mortem collected tissue samples.
The children under 1 year of age had died of sudden infant death syndrome (SIDS). In studies of SIDS, no differences have been found in the mucosal immunology of the respiratory tract.35 It is therefore unlikely that the data are biased for SIDS.
Expression of markers and correlation
Serial single staining was performed and revealed correlations between the expression of DC-SIGN and different co-stimulatory and adhesion molecules. A similar method calculating co-expression of surface molecules of DC has been used for the skin,6 and the technique of sequential fluorescent overlays has also been reported previously.27 The marker DC-SIGN recognises mature and immature mucosal DC36 but is also expressed by alveolar macrophages, interestingly only after birth.11 The functional relevance has recently been shown in a pig model where trachea and gut tissue were investigated.32 The co-expression of CD80/86 of DC indicated an activated or maturing phenotype. In contrast, using cells extracted from human lung lobes, the phenotype of DC has been reported to be mainly immature.37 In the rat model it has been shown that T cells are not only activated in the draining lymph nodes, but also transiently in the lung itself after reception of cognate signals from local memory T cells.38 Such interactions have not been seen in human bronchial mucosa to date. It might be that recurrent infections and the continuous antigenic stimulation even in the “normal” human respiratory tract lead to cognate signals for DC activation. Data from the rat trachea and larynx have shown the influence of localisation and infectious stimuli on colonisation with DC.39 It has recently been reported that the presence of DC in the human respiratory mucosa is modified by infections.40
It is difficult to study such dynamic interactions for samples of human tracheobronchial and peripheral lung tissues. It remains unclear whether different compartments of the respiratory tract could be compared because of major differences in anatomy, function, and blood supply.41 Future studies should focus on the morphology and function of epithelial DC since it has been shown in the gut that dendrites of DC are able to open tight junctions and search the lumen to pick up antigen.42,43
Influence of age
We have previously reported that the density of DC, determined by HLA-DR staining and morphology in paraffin sections, is different in infants under the age of 1 year than in older individuals.18 This is similar to data in the rat trachea in which the density of DC reached adult levels 2 weeks after birth.44 In the present study the patients were also divided into two age groups (younger or older than 1 year of age), and age dependent differences were found at certain topographical sites. The probable reason for the lower cell numbers in children under the age of 1 year is that they have not yet encountered many antigens and their immune system is beginning to develop. The ratios of cell densities in the lamina propria and submucosa were the same in both groups for all markers. This indicates that the cellular distribution between the layers is already established early in life.
Influence of localisation
The decline in cell numbers from the ventral to the dorsal sites was not evident in infants under 1 year of age. The higher cell densities in the ventral tracheal mucosa of older patients indicate that this is immunologically the most active site. One explanation for this could be a higher impact of antigens and microorganisms caused by non-laminar airflow at the tracheal ring, while at the posterior membranous wall the airflow is laminar. Interestingly, the opposite finding has been described for the rat trachea in the steady state, where Schon-Hegrad et al24 argue that differences in the vascularisation might account for the higher number of DC at the dorsal tracheal site. Compartmentalisation of functional DC subsets has recently been shown in the mouse.45 Moreover, a few substances which target DC in pulmonary allergic inflammation have become available—for example, a new low molecular weight immunomodulator has been found to inhibit DC-mediated T helper cell activation in a mouse model of allergic lung inflammation.46 With such therapeutic perspectives, information is urgently needed on DC subsets in the human lung.
To our knowledge, this is the first study of DC in the human tracheal mucosa in a reasonably large number of tissue samples. The relative distribution in the mucosal layers is already established during the early phase of life. The human tracheal mucosa contains many molecules needed for local immune interactions. The finding of site specific differences between the ventral and dorsal site of the trachea has not been reported previously and should be taken into consideration in future investigations. The increasing density of mucosal DC with age at the ventral trachea could reflect maturation of the mucosal immune system.
Acknowledgments
The authors thank Karin Westermann, Gerhard Bargsten, and Henning Weigt for technical help, and Sheila Fryk for correcting the English.
REFERENCES
Footnotes
-
Published Online First 7 August 2006
-
TT and VCdV contributed equally to the paper.
-
This study was supported by a grant from the Deutsche Forschungsgemeinschaft (SFB 587/B5).
-
Competing interests: none.
Linked Articles
- Airwaves