Article Text

Abstract
Cilia are organelles present on almost every cell within the body. In the respiratory tract, motile cilia line the epithelial surface and beat in a coordinated fashion to clear mucus from the airways. Mucociliary dysfunction is implicated in a number of lung diseases including cystic fibrosis, chronic obstructive pulmonary disease, bronchiectasis and primary ciliary dyskinesia. Recent microscopy advances such as electron tomography have developed our understanding of the structure and function of these important organelles. In this review we discuss electron tomography and its relevance to respiratory cilia.
- Airway Epithelium
- Histology/Cytology
- Imaging/CT MRI etc
- Bronchiectasis
- Paediatric Lung Disaese
- Rare lung diseases
Statistics from Altmetric.com
- Airway Epithelium
- Histology/Cytology
- Imaging/CT MRI etc
- Bronchiectasis
- Paediatric Lung Disaese
- Rare lung diseases
Introduction to electron tomography
Dual axis electron tomography is an advanced electron microscopy technique used for the visualisation of structures in three dimensions. A series of transmission electron microscopy images are acquired by tilting the specimen stage at regular increments (typically angles range from −70° to +70°) around two perpendicular axes. Images from both tilt series are then aligned into a single three-dimensional high-resolution projection. If a structural feature is repeated within a tomogram, it can be enhanced through sub-tomographic averaging; a technique in which software extracts the chosen common features and makes comparison by cross-correlation. Within the respiratory epithelium the cilium makes a tempting target for this technique, given the repetitive nature of its axonemal structure. Tomography combined with sub-tomographic averaging can achieve a resolution of approximately 5 nm, sufficient to distinguish molecular complexes. In addition, the three-dimensional information can give a novel insight into the conformation of structures and their interactions.
Ciliary ultrastructure
The fundamental structure of the human respiratory ciliary axoneme, as shown in figure 1, is highly conserved among eukaryotic organisms and consists of nine peripheral microtubular doublets arranged radially around a central microtubular pair. Connecting the central pair and each microtubular doublet are a series of radial spokes, and between doublets are nexin links. On each outer microtubular doublet are motor proteins known as the inner and outer dynein arms which drive ciliary beating through ATPase activity. This whole structure extends from the cell and is surrounded by a ciliary membrane. Longitudinally, the key components of the axoneme repeat in units of approximately 96 nm which combine to produce a full length respiratory cilium of 6–7 µm.
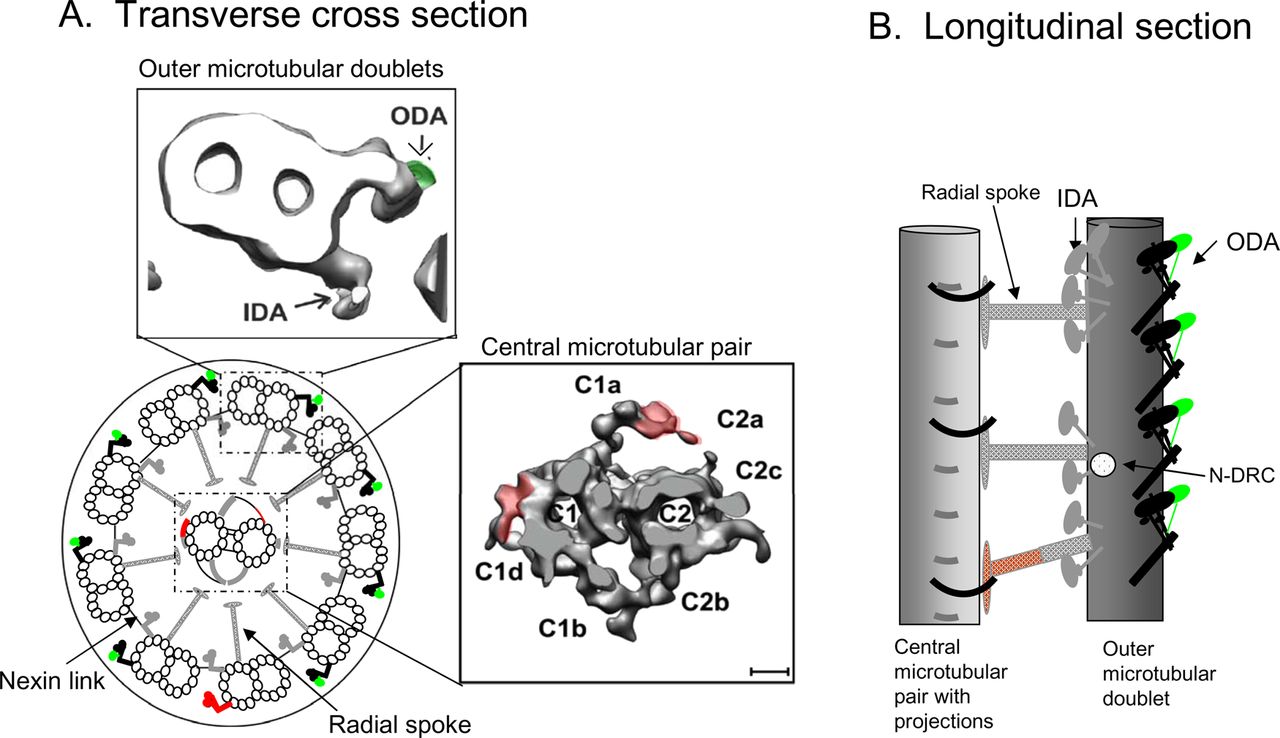
{kind=link}
(A) Transverse cross-section and (B) longitudinal section showing the structure of the human respiratory ciliary axoneme. IDA, inner dynein arm; N-DRC, dynein regulatory complex; ODA, outer dynein arm. Red shows structures absent or altered in Chlamydomonas flagella. Green depicts structures present in Chlamydomonas flagella but absent in human respiratory cilia.
Insights into cilia ultrastructure by electron tomography
Electron tomography on flagellated organisms such as the blue-green alga Chlamydomonas reinhardtii has identified new structural and functional relationships within the axoneme. For example, since being described in the 1960s, the function of the nexin link remained elusive until the elegant use of cryo-electron tomography in Chlamydomonas identified that it is part of a larger dynein regulatory complex (N-DRC) which plays a pivotal role in the regulation of beat frequency.1 Subsequently, it was hypothesised that signals controlling flagella movement are transmitted to the N-DRC through the radial spokes. Electron tomography conducted on wild type and a series of mutant Chlamydomonas identified a calmodulin and spoke-associated complex required for flagella movement. Its structural location makes it likely to be responsible for the transmission of signals to the N-DRC and ultimately to the dynein motors.2 Because of the highly conserved nature of the axonemal structure, findings from flagellated model organisms such as those described can usually be translated directly to human cilia. However, species differences have been highlighted. For example, cryo-electron tomography demonstrated that the protozoa Tetrahymenia spp has a third distinctly different radial spoke compared with Chlamydomonas.3 The third spoke is also present in the human respiratory cilium, which has recently been tomographically modelled.4 Figure 1 shows in detail the central microtubular pair and an outer microtubular doublet from the human model compared with the well-characterised Chlamydomonas. This model axoneme highlights features of human cilia such as the two heavy chain dyneins on the outer dynein arm compared with three in Chlamydomonas, the projections that surround the central microtubular doublet and the three distinct radial spokes that radiate around the central microtubular pair in a helical arrangement.
Practical applications of electron tomography in respiratory disease
In the genetic condition primary ciliary dyskinesia (PCD), respiratory cilia dysfunction results in chronic wet cough, repeated chest infections, rhinosinusitis and otitis media. If PCD is not diagnosed promptly, permanent lung damage can occur in the form of bronchiectasis. A number of established structural defects are known to cause PCD, the majority of which are characterised by the absence of one or both of the dynein arms. By standard transmission electron microscopy, however, approximately 15% of patients have an apparently normal ultrastructure. Confidence in diagnosis of these patients is low, and patients often undergo several repeat nasal biopsies in attempts to establish a diagnosis.
Electron tomography can be used to identify the structural defects in some of these cases. One known genetic cause of PCD, defined by the absence of the outer dynein arm heavy chain DNAH11, is associated with ‘normal ultrastructure’ when visualised by standard transmission electron microscopy.5 The individual outer dynein arm heavy chains can be resolved using electron tomography, making detection of DNAH11 mutations possible (unpublished data). In addition, electron tomography, genetics and molecular biology used in combination have recently identified the absence of Hydin as another cause of PCD. Electron tomography showed that projections associated with the central pair apparatus (C2b and C2c) are absent in patients with a Hydin defect, suggesting a location for Hydin in the normal axoneme.6 As with DNAH11 mutations, the cilia from patients with Hydin appear to be structurally normal when viewed with standard transmission electron microscopy, further highlighting the benefits of using electron tomography in cases of PCD that are difficult to diagnose.
The limitations of electron tomography concern microscopic resolution, data inaccessible through tilting of the sample and the speed of data processing. However, with rapidly increasing processing power, high voltage electron microscopes allowing penetration of thicker samples and high sensitivity recording devices, we anticipate the biggest obstacle of the future will be the interpretation of these large datasets. While electron tomography is used within the NHS in assessment of viral structure7 and will be useful in specialist cases of PCD diagnosis, we predict the most important role will be its use in combination with molecular and genetic techniques in unravelling the complexity of these small yet significant structures.
Acknowledgments
The authors wish to thank members of the Royal Brompton Hospital Electron Microscopy Unit and the NHS Advisory Group for National Specialised Services (AGNSS) for their support.
Footnotes
Contributors AS and CH were both involved in researching, planning and drafting this article. Both authors approved the final version.
Funding None.
Competing interests None.
Provenance and peer review Commissioned; internally peer reviewed.
Linked Articles
- Airwaves