Article Text

Abstract
Rationale Muscle atrophy confers a poor prognosis in patients with chronic obstructive pulmonary disease (COPD), yet the molecular pathways responsible are poorly characterised. Muscle-specific microRNAs and serum response factor (SRF) are important regulators of muscle phenotype that contribute to a feedback system to regulate muscle gene expression. The role of these factors in the skeletal muscle dysfunction that accompanies COPD is unknown.
Methods 31 patients with COPD and 14 healthy age-matched controls underwent lung and quadriceps function assessments, measurement of daily activity and a percutaneous quadriceps muscle biopsy. The expression of muscle-specific microRNAs, myosin heavy chains and components of the serum response factor signalling pathway were determined by qPCR.
Results A reduction in expression of miR-1 (2.5-fold, p=0.01) and the myocardin-related transcription factors (MRTFs) A and B was observed in patients compared with controls (MRTF-A mRNA: twofold, p=0.028; MRTF-B mRNA: fourfold, p=0.011). miR-1 expression was associated with smoking history, lung function, fat-free mass index, 6 min walk distance and percentage of type 1 fibres. miR-133 and miR-206 were negatively correlated with daily physical activity. Insulin-like growth factor 1 mRNA was increased in the patients and miR-1 was negatively correlated with phosphorylation of the kinase Akt. Furthermore, the protein levels of histone deacetylase 4, another miR-1 target, were increased in the patients.
Conclusions Downregulation of the activity of the MRTF-SRF axis and the expression of muscle-specific microRNAs, particularly miR-1, may contribute to COPD-associated skeletal muscle dysfunction.
- Muscle phenotype
- microRNA
- serum response factor
- COPD
- COPD mechanisms
- respiratory muscles
- sleep apnoea
- COPD exacerbations
- long term oxygen therapy (LTOT)
- lung physiology
- non invasive ventilation
- pulmonary rehabilitation
- allergic lung disease
- assisted ventilation
- asthma
- lung cancer
- non-small cell lung cancer
- sarcoidosis
- small cell lung cancer
- tuberculosis
- COPD pathology
- lung volume reduction surgery
- respiratory measurement
- exercise
- systemic disease and lungs
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/2.0/ and http://creativecommons.org/licenses/by-nc/2.0/legalcode.
Statistics from Altmetric.com
- Muscle phenotype
- microRNA
- serum response factor
- COPD
- COPD mechanisms
- respiratory muscles
- sleep apnoea
- COPD exacerbations
- long term oxygen therapy (LTOT)
- lung physiology
- non invasive ventilation
- pulmonary rehabilitation
- allergic lung disease
- assisted ventilation
- asthma
- lung cancer
- non-small cell lung cancer
- sarcoidosis
- small cell lung cancer
- tuberculosis
- COPD pathology
- lung volume reduction surgery
- respiratory measurement
- exercise
- systemic disease and lungs
Key messages
What is the question?
How are changes in microRNA expression and serum response factor (SRF) activity associated with changes in skeletal muscle phenotype in patients with chronic obstructive pulmonary disease (COPD)?
What is the key point?
The expression of the myocardin-related transcription factors (activators of SRF) and of miR-1 (an SRF target) is reduced in the quadriceps of patients with COPD.
Why read on?
The study identifies the SRF/miR-1 axis as an important contributor to muscle phenotype in patients with COPD.
Introduction
Skeletal muscle dysfunction in the locomotor muscles is an important systemic complication of chronic obstructive pulmonary disease (COPD). Quadriceps weakness and wasting predict mortality among patients with moderate to severe COPD independently of lung function1 2 and are common complications.3 Characteristically, the vastus lateralis of patients with COPD exhibits a shift towards a predominance of type IIA fibres and an increase in the proportion of type IIX fibres associated with a marked reduction in exercise capacity.4 5 In addition to these changes, fibre atrophy occurs and is particularly marked in type IIX fibres. However, the mechanisms that control these phenotypic changes have not been fully defined.
Recent data have implicated microRNAs (miRNAs, small non-coding RNAs that reduce mRNA half-life and translation) in the regulation of the skeletal muscle phenotype.6 Several miRNAs are highly expressed in skeletal muscle and regulate muscle phenotype. miR-1 and miR-206 promote myotube formation7 and miR-206 is associated with regeneration after nerve injury.8 Genetic knockout of miR-499 or miR208-b reduces the proportion of type I fibres whereas overexpression of miR-499 increases type I fibres and endurance in mice.9 10 Conversely, miR-133 expression inhibits myotube formation and promotes proliferation.11
Analysis of miRNA expression in primary skeletal myopathies identified changes in miRNAs that target conserved pathways, but the expression of individual miRNAs varied.12 Similarly, the profile of miRNAs is altered in the skeletal muscle of patients with insulin resistance.13 miRNA profiles are also markedly affected by activity, and studies in humans have shown altered miRNA profiles associated with endurance exercise training.14 As inactivity may be a major contributor to skeletal muscle dysfunction in COPD, changes in miRNA expression associated with inactivity may contribute to the phenotype. Studies of inactivity resulting from denervation,15 nerve entrapment16 or space flight17 all show initial reductions in miR-1, but whether miR-1 levels remain suppressed varies. Interestingly, miR-1 did not change in response to resistance training in humans but there was altered expression of miRs associated with the mTOR pathway.18
The myocardin-related transcription factor (MRTF)-serum response factor (SRF) axis regulates the expression of miR-119 and other muscle-specific miRNAs.20 Changes in the expression and localisation of SRF and the SRF co-activators, MRTF-A and MRTF-B, in response to physical activity and ageing have all been reported.21–23 In addition to miRNA expression, MRTF/SRF activity is important in regulating MHC expression. SRF activity is more important in activating expression of MHCI than MHCIIa and MHCIIx, implying a role for SRF in fibre-type control.24 The miRNAs feed back to regulate MRTF/SRF and other factors that control muscle cell phenotype such as myocyte enhancer factor-2 (MEF-2), and proliferation including insulin-like growth factor 1 (IGF-1)25 and the myostatin signalling pathway.26 27 Together these MRTF/SRF-miRNA interactions provide the system shown in figure 1.

Relationships between miRNAs and selected target genes in muscle. The relationship between miRNAs and the RNAs that encode proteins is shown. miRNA targets are either predicted using targetscan (http://www.targetscan.org/) or verified by experiment. HDAC4, histone deacetylase 4; IGF-1, insulin-like growth factor 1; MRTF, myocardin-related transcription factor; SRF, serum response factor; Act IIB receptor, activin type IIB receptor.
We hypothesised that reduced physical activity commonly observed in patients with COPD would suppress SRF activity and reduce expression of miR-1 and other SRF-dependent miRNAs along with altered expression of miRNA targets in comparison with healthy controls. We therefore analysed muscle-specific miRNA expression, the expression of miR-145 (an miRNA associated with SRF activity in smooth muscle) and miR-181 (an miRNA altered by activity but not known to be SRF-dependent28) along with the expression of components of the SRF pathway in quadriceps biopsies from patients with COPD and age-matched controls. Some of the results presented here have been reported in abstract form.29
Methods
Subjects
Thirty-one patients with COPD according to the Global Initiative in Obstructive Lung Disease guidelines 200430 were enrolled from clinics at the Royal Brompton Hospital. Fourteen healthy age-matched controls were recruited by advertisement. Patient exclusion criteria and ethical approval are detailed in the online supplement.
Physiological measurements
General assessment
Lung volume, carbon monoxide transfer, blood gas tension and fat-free mass index (FFMI) were measured as described in the online supplement.
Muscle assessments
Quadriceps strength was determined by measuring maximal voluntary contraction (MVC) and unpotentiated twitch quadriceps force (TwQ). Exercise performance was measured using a 6 min walk (6MW) test and daily physical activity was measured using a tri-axial accelerometer. Detailed descriptions of the physiological methods are given in the online supplement.
Assessment of mRNA, miRNA and protein levels
Messenger RNA was extracted and quantified as described previously31 and detailed in the online supplement. MicroRNA expression was analysed in trizol extracted RNA using the Ncode SYBR green miRNA-qRT-PCR kit (Invitrogen, Paisley, UK) as detailed in the online supplement. Protein was analysed by luminex or western blotting as described in the online supplement. The amount of material available did not allow us to measure all values in all subjects.
Assessment of MRTF activity
The activity of MRTFs on the miR-1 promoters was determined as described in the online supplement.
Immunofluorescence analysis
Fibre size, fibre proportion and SRF localisation were determined by immunofluorescence as described in the online supplement.
Statistical analysis
Unsupervised principal components analysis (PCA) and unsupervised hierarchical clustering were performed in Aabel (Gigawiz). The variables for hierarchical clustering were scaled to unit variance and distances calculated based on correlation coefficient similarity. Groups were defined based on centroid linkage. Log transformation of the miRNA data produced a normal distribution for all the miRNAs. Correlation analysis was performed using the Pearson correlation coefficient. To interpret the correlation data, p<0.05 was taken as an indicator of association to allow us to investigate likely relationships within the data, but none of the relationships were significant if applying a Bonferroni correction.
Differences between patients and controls were calculated by the Student t test for normally distributed data and by the Mann–Whitney U test for data that did not fit a normal distribution. The test used for each analysis is denoted by T (t-test) and MW (Mann–Whitney U) in parentheses. The statistical significance of differences was taken at p<0.05.
Results
Patient characteristics and muscle phenotype
As expected, the patients had significant lung function impairment and reduced arterial blood oxygen tensions, consistent with a diagnosis of COPD. However, there was no significant difference in age, weight, body mass index (BMI) or arterial blood carbon dioxide tensions between groups and the groups were matched for gender (p=0.74, Fisher exact test). Patients had a significantly reduced FFMI, quadriceps force and locomotion time compared with controls (table 1).
Clinical characteristics of study subjects
The patients had a marked difference in the size and proportion of muscle fibres compared with the controls (see figure S1 in online supplement). The proportion of type I fibres in the patients was reduced (from a median of 52 (IQR 39–62) to 25 (IQR 17–30)) and the proportion of type II fibres was increased (from mean±SD 47±14 to 69±15, table 1). The proportion of hybrid type I/type II fibres was also increased in patients (from a median of 1.5 (IQR 0–3) to 3 (IQR 0–11), table 1). The fibres of the patients were smaller than those of the controls with a significant reduction in the size of the type I and type IIX fibres (table 1).
Patients had reduced MHCI mRNA compared with controls (table 1 and figure S2 in online supplement). MHCI mRNA was directly correlated with forced expiratory volume in 1 s (FEV1) percentage predicted and inversely correlated with smoking history across both patients and controls (see figure S2 in online supplement). MHCIIa expression was not correlated with smoking history or FEV1 (not shown). MHCI mRNA was positively correlated with exercise performance (6MW, figure S3 in online supplement) but was not associated with strength. MHCIIa was inversely correlated with 6MW (figure S3 in online supplement) across all data as well as within the patient group alone, but also showed no association with strength. There was no significant difference in α-actin expression between the groups (data not shown).
miRNA expression in quadriceps of patients with COPD
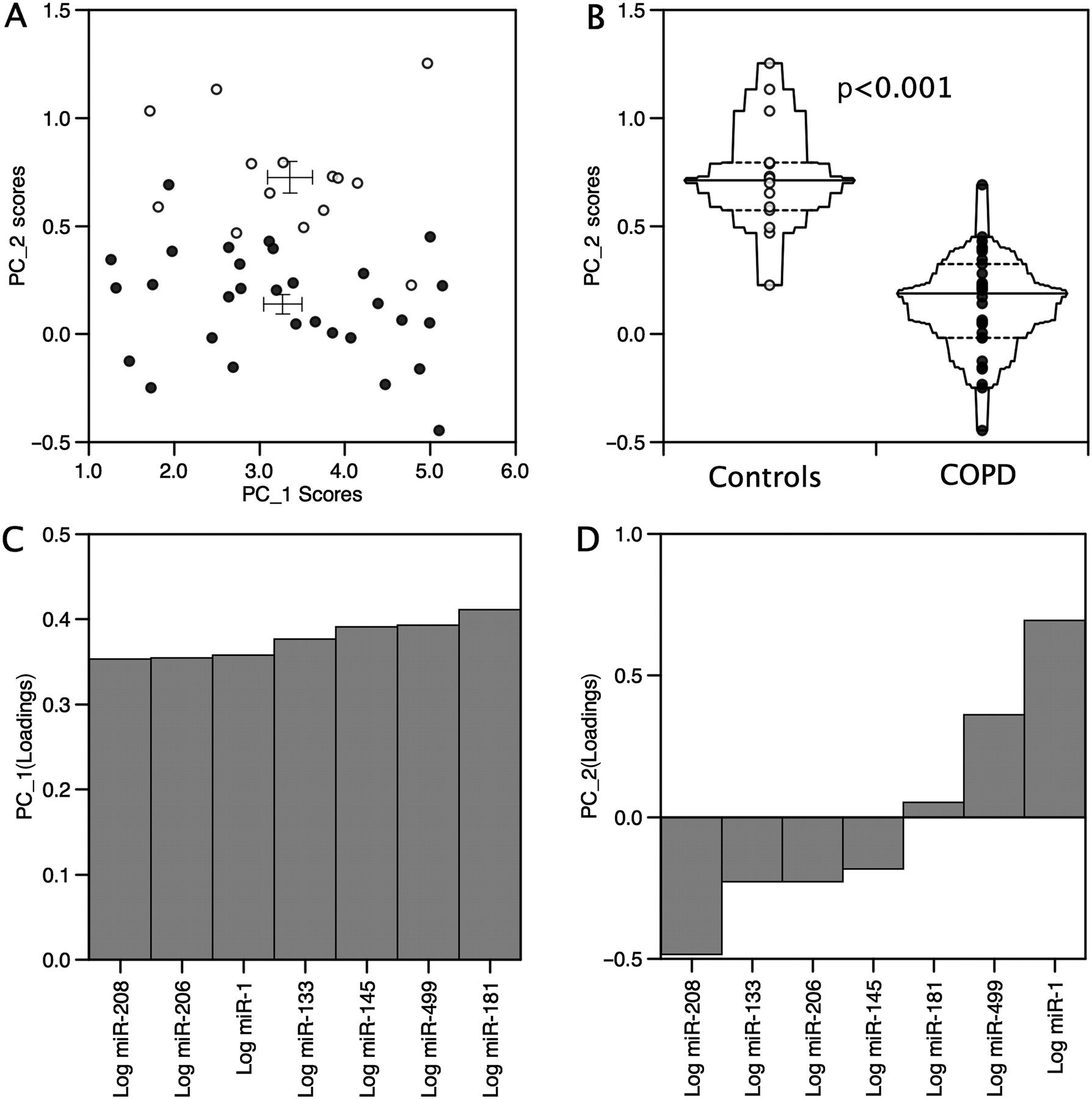
Multivariate statistical analyses (eg, hierarchical clustering and PCA) offer a powerful unsupervised approach for the identification of structure and relationships between samples, allowing for the visualisation of natural groupings in data. Hierarchical clustering (figure 2) and PCA of miRNA expression identified a distinct profile for the patients compared with the controls (PCA, p<0.0001, figure 3B), primarily associated with an approximate 2.5-fold reduction in the expression of miR-1 (p=0.01(T), figure 3D and figure 4) in patients compared with controls, but there was no change in the expression of miR-133 (figure 4). Differences in the expression of miR-208 and miR-499 contributed to the observed difference in the miRNA expression pattern (figure 3D), although taken alone these did not reach statistical significance (miR-208, p=0.099, miR-499, p=0.142(T), figure 4). Hierarchical clustering of the expression patterns of individual miRNAs within the samples showed that miR-1, miR-133 and miR-206 formed a cluster consistent with their expression as bicistronic RNAs from the miR-1/miR-133 and miR-206/miR-133 genes, providing validation of our methodology.

miRNA expression is markedly different in patients with chronic obstructive pulmonary disease (COPD) compared with matched controls. The expression of miRNAs was determined by qPCR and normalised to the expression of 5S RNA in the same sample, as described in the Methods section. The heat map shows the expression of miRNAs in each sample organised by hierarchical clustering in which the expression of each miRNA was given equal weighting. This analysis shows clear separation of the majority of patients (closed circles) from the control group (open circles).

Principal component analysis of the miRNA expression pattern in patients with chronic obstructive disease (COPD) and controls. (A) Scatter plot comparing PC1 and PC2 for each sample. (B) Principal component analysis of miRNA expression in patients and controls shows a significant difference in the pattern of miRNA expression between the two groups as a difference in PC2 (p<0.0001). (C) Loading plots for PC1 showing equal contribution of the miRNAs to this component. (D) Loading plots for principal component analysis identifies miR-1, miR-208 and miR-499 as the major contributors to the separation of the two groups. Patients are shown as closed circles and controls are shown as open circles.
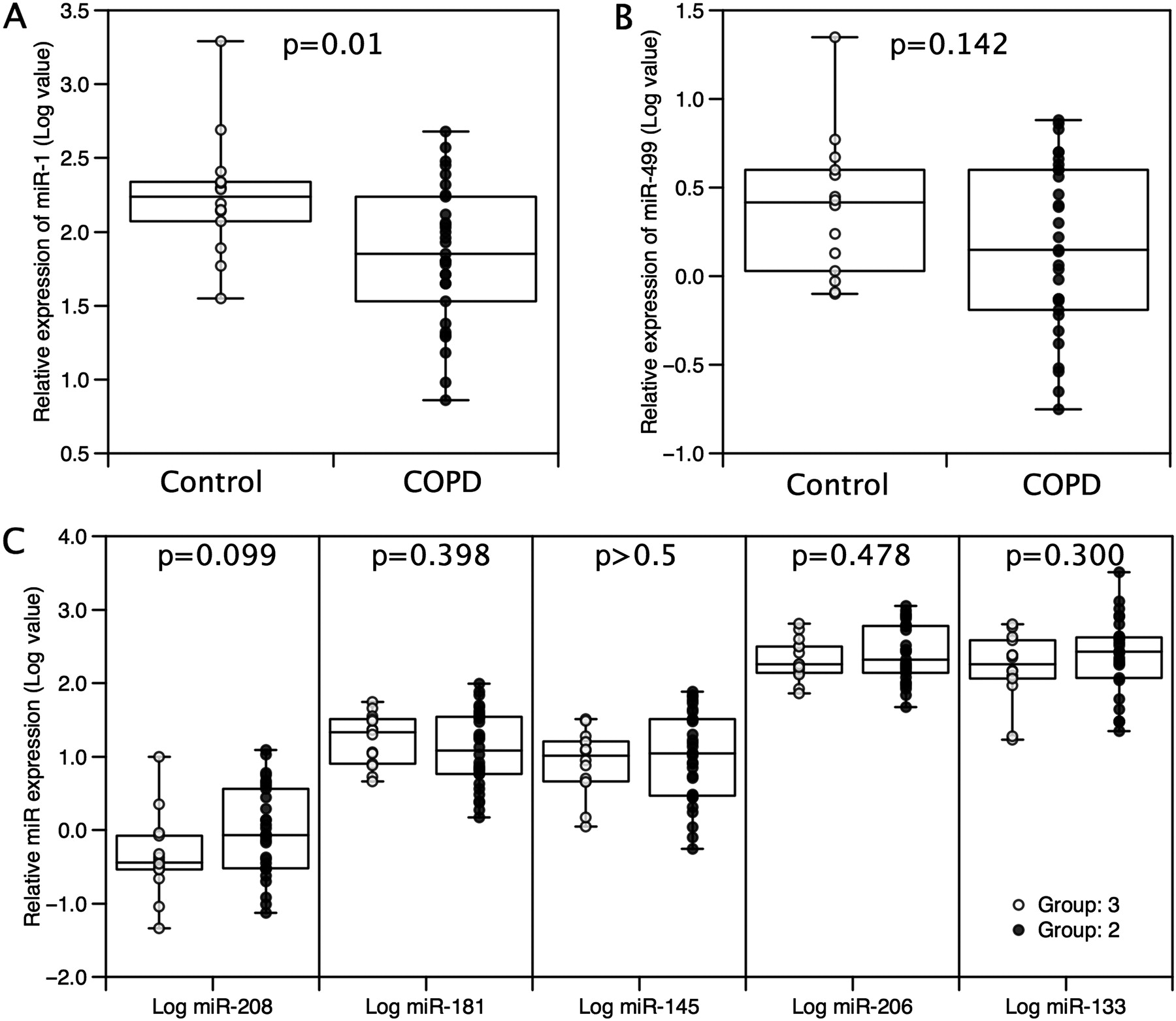
Expression of microRNAs in patients with chronic obstructive pulmonary disease (COPD) and controls. Expression of miRNAs was determined by qPCR and normalised to the expression of the 5S RNA in the same sample as described in the Methods section. The expression of miR-1 (A) was lower in patients than controls but there was no significant difference in the expression of miR-499 (B) or the other miRNAs (C) analysed. Data are presented as log normalised expression with the box showing median and IQR, error bars to maximum and minimum points. Patients are shown as closed circles and controls are shown as open circles. Statistical significance was calculated by t test.
Comparison of the expression of the miRNAs with the physiological data showed that miR-1 was negatively correlated with smoking history (r=−0.39, p=0.007) but positively associated with FEV1 (r=0.34, p=0.022), FFMI (r=0.33, p=0.025), 6MW (r=0.33, p=0.026) and MVC/BMI (r=0.29, p=0.049). Like miR-1, miR-499 was positively associated with FFMI (r=0.37, p=0.012) and negatively with smoking history (r=−0.33, p=0.027). miR-133 and miR-206 were negatively associated with physical activity (movement intensity; miR-133: r=−0.30, p=0.057; miR-206: r=−0.35, p=0.023 and locomotion time; miR-133: r=−0.35, p=0.027; miR-206: r=−0.34, p=0.029). The association of the miRNAs with different physiological characteristics is shown in figure 5A. Associations with p<0.05 were only present if both the control and patient groups were combined; this feature may result from a lack of power but also raises the possibility that disease is an important component of the difference.
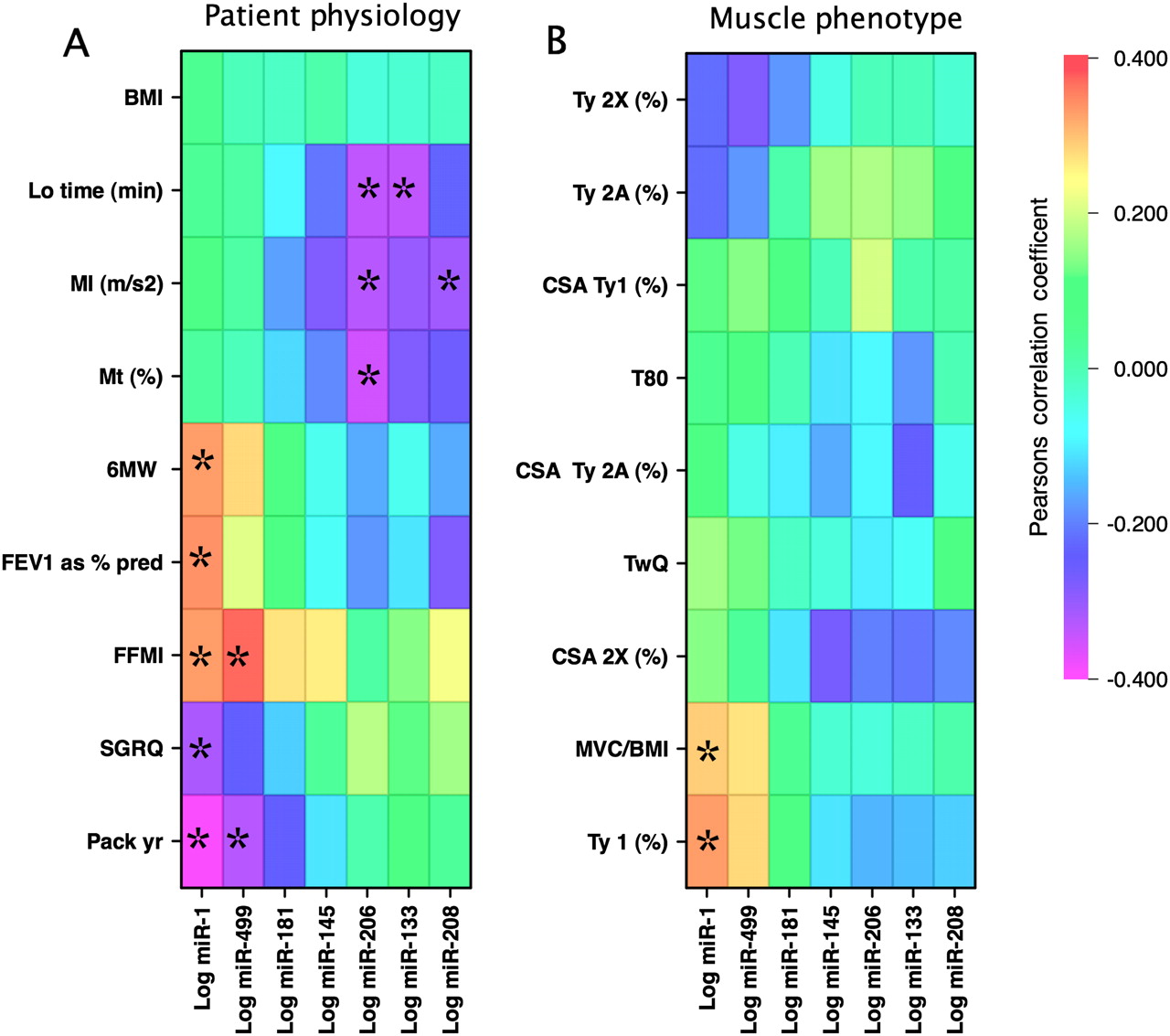
Pearson correlation matrices for miRNAs with physiological characteristics and muscle fibre parameters in the cohort. MicroRNA expression was correlated with non-muscle physiological characteristics for all the samples (A) or with the muscle-specific physiological and histochemical parameters (B). The direction and intensity of the correlation is colour-coded according to the bar. The miRNAs are organised in the order of their contribution to PC2 as determined by principal components analysis. Correlations reaching a statistical significance where p<0.05 are indicated by * and the p values are given in table S1 in the online supplement. Characteristics were ordered by hierarchical clustering (figure S4 in online supplement). BMI, body mass index; CSA, cross-sectional area; FFMI, fat-free mass index; FEV1, forced expiratory volume in 1 s; Lo, locomotion; MVC, maximal voluntary contraction; MI, movement intensity; Mt, movement time; 6MW, 6 min walk test; SGRQ, St George Respiratory Questionnaire; TwQ, twitch force in the quadriceps; Ty, type.
Comparison of miRNA expression with the muscle characteristics showed that miR-1 was positively correlated with the percentage of type I fibres (r=0.33, p=0.029) and miR-499 showed a trend towards association with percentage of type I fibres (r=0.28, p=0.063). None of the other miRNAs was associated with muscle fibre characteristics (figure 5B).
Expression of SRF, MRTF-A, MRTF-B
To investigate the SRF system we analysed the expression of SRF and two SRF co-activators (MRTF-A and MRTF-B) by qPCR. Mean SRF mRNA was not significantly different between the groups (COPD 3.42 (IQR 2.49–3.63) AU, n=19; controls 3.31 (IQR 3.22–3.89) AU, n=16, p=0.185 (MW), figure 6A). MRTF-A and MRTF-B mRNA levels were significantly lower in patients (3.73 (IQR 3.48–3.87) AU and 3.94±0.394 AU respectively, n=25) than in controls (3.85 (IQR 372–4.22) AU, p=0.028 (MW), figure 6B and 4.42±0.634 AU, p=0.011 (T) respectively, n=15, figure 6C). MRTF-A expression was correlated with FEV1 (% predicted, r= 0.46, p=0.027) and MRTF-B was correlated with smoking history (r=−0.576, p=0.006, not shown). Again these associations were not present in individual groups. To tie this reduction in MRTF expression to miR-1, we showed that MRTFs could activate the miR-1 promoters (figure 6D and supplementary results).

Effect of myocardin-related transcription factors (MRTFs) on miR-1 promoter activity and the expression of MRTF and serum response factor (SRF) in the quadriceps muscle of patients with chronic obstructive pulmonary disease (COPD). Expression of MRTF-A (A), MRTF-B (B) and SRF (C) mRNA in the quadriceps of patients and healthy age-matched controls was quantified by real-time PCR and normalised to the expression of RPLPO. Data are presented as log normalised expression with the box showing median and IQR, error bars to maximum and minimum points. The expression of MRTF-A and MRTF-B was suppressed in the patients compared with controls but SRF expression did not differ significantly between groups (AU, arbitrary units). (D) C2C12 cells were transfected with miR-1-1, miR-1-2 promoter reporter vectors as described in the online supplement in the presence or absence of expression vectors for MRTF-A and MRTF-B. Luciferase activity was determined 24 h later. MRTF-A and MRTF-B increased the activity of the miR-1-1 and miR-1-2 promoters (p<0.001) but did not increase the activity of the delta enhancer promoter. Data are presented as mean±SEM. Patients are shown as closed circles and controls are shown as open circles. Statistical significance for the mRNA expression was calculated by the Mann–Whitney U test.
SRF activity is also regulated by the localisation of the protein. We therefore determined the localisation of SRF in muscle sections from a subset of patients with COPD and controls. In the controls, SRF was readily detectable in the nuclei of all of the samples analysed. However, in the patients there was a reduction in nuclear staining and the appearance of nuclei with perinuclear SRF staining (figure 7).

Serum response factor (SRF) localisation is altered in patients with chronic obstructive pulmonary disease (COPD). The localisation of SRF was determined in sections of quadriceps muscle from patients with COPD and controls by immunofluorescence as described in the online supplement. Arrowheads show the localisation of nuclei and the inset shows perinuclear staining for SRF.
Reduced expression of targets of miR-1 in patients with COPD
miR-1 targets include IGF-1 and the IGF-1 receptor,25 indicating that it should modify the activity of the IGF-1 pathway, and we have found that phosphorylation of the kinase Akt (also known as protein kinase B) is inversely proportional to activity in our patient group.32 We therefore analysed the expression of IGF-1 and the activity of the IGF-1 pathway by examining the phosphorylation of Akt in patients and controls. IGF-1 mRNA was increased in patients (figure 8A) and miR-1 expression was inversely proportional to the phospho-Akt/total Akt ratio (figure 8B, r=−0.43, p=0.022). A trend towards this association was present in the patients alone (r=−0.46, p=0.053). There were no other associations of the phospho-Akt/total Akt ratio with any of the other miRNAs.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Altered insulin-like growth factor 1 (IGF-1) and histone deacetylase 4 (HDAC4) in patients with chronic obstructive pulmonary disease (COPD). IGF-1 (A) and HDAC4 (C) mRNA were quantified by real-time PCR as described in the online supplement. IGF-1 mRNA was increased whereas HDAC4 mRNA was unchanged in patients relative to controls. Data are presented as log normalised expression with the box showing median and IQR, error bars to maximum and minimum points. Akt levels and phospho-Akt levels were determined by lunminex assay in muscle homogenates prepared as described in the online supplement. miRNA expression was quantified by qPCR and normalised to the expression of 5S RNA. Pearson correlation coefficients showed that miR-1 (B) was correlated with the ratio of phospho-Akt to total Akt. Patients are shown as open circles and controls are shown as closed circles. (D) HDAC4 protein levels were detected by western blotting (inset) quantified by densitometry and normalised to total protein loaded onto the blot determined by Ponceau Red staining. HDAC4 protein levels were increased in patients compared with controls. Data are presented as normalised protein levels with the box showing median and IQR, error bars to maximum and minimum points. Statistical significance for mRNA and protein levels was determined by the Mann–Whitney U test. Due to limitations on specimen size, we were unable to measure HDAC4 protein and miRNA in the same sample set.
We also determined the expression of histone deacetylase 4 (HDAC4) mRNA and protein in an additional set of patients with COPD and controls. This analysis showed that, while there was no difference in the expression of HDAC4 mRNA between patients and controls (figure 8C), HDAC4 protein was higher in patients with COPD than in controls (figure 8D).
Discussion
This study shows that the profile of miRNAs in the quadriceps of patients with COPD differs from that in controls. The data also suggest that MRTF/SRF activity is downregulated in patients. Together with previous animal data showing a role for SRF and the miRNAs in the control of muscle phenotype, these data support the hypothesis that altered miRNA expression and reduced SRF activity contribute to COPD-associated muscle dysfunction.
Significance of the findings
The most striking finding is the reduction in the expression of miR-1 in patients compared with controls. miR-1 expression is increased by SRF,19 and we show that it is increased in an SRF-dependent manner by the MRTFs. The reduced expression of MRTFs, as well as altered SRF localization, suggest that reduced activity of the MRTF/SRF axis contributes to the reduction in MHCI and miR-1 expression. Our data also indicate functional consequences of reduced miR-1 with increased expression of IGF-1, an inverse correlation of miR-1 with Akt phosphorylation and increased HDAC4 protein in the patients. Whether the observed increase in IGF-1 mRNA is a direct response to reduced miR-1 is not clear, as miRNAs are suggested to inhibit translation in muscle rather than increase RNA degradation.13 Consistent with this suggestion, we found that patients had normal HDAC4 mRNA levels but increased HDAC4 protein. The interaction of miR-1 with IGF-1 signalling and HDAC4 have been documented previously.25 33–35 These observations raise two points regarding the role of the SRF/miR-1 axis in muscle, which are discussed separately.
miR-1 in muscle biology
miR-1 has an accepted role in skeletal muscle differentiation. One likely effect of the reduced miR-1 observed is the increase in HDAC4 protein. HDAC4 inhibits the activity of MEF-2 and SRF, both of which are important regulators of MHCI expression, providing a mechanism by which the reduction in miR-1 may contribute to reduced MHCI and fibre shift. The fact that miR-1 is an SRF target provides a positive feedback loop that may contribute to the downward spiral in muscle phenotype. HDAC4 also suppresses the expression of follistatin: reduced miR-1 would therefore increase the activity of the myostatin pathway and promote muscle wasting.36 These effects of miR-1 suggest a role in the maintenance of muscle phenotype and, consistent with these suggestions, miR-1 expression was inversely proportional to the percentage of type I muscle fibres. The reduction in miR-1 in patients, who have a lower level of activity than controls, is different from the response to endurance training in young adults.18 miR-1 has also been suggested to inhibit the calcineurin pathway, thereby suppressing activity-dependent MEF-2 activation.37 The activity of miRNAs is therefore likely to be cell and context dependent.
miR-1 also inhibits the expression of IGF-1 and, consistent with previous reports,25 we found an inverse correlation between miR-1 and IGF-1 signalling. A similar increase in IGF-1 mRNA has previously been observed by mRNA profiling in COPD.38 The increase in IGF-1 mRNA in patients likely to show muscle wasting seems paradoxical. However, other studies have also shown increased activity of the hypertrophy signalling pathway in patients with COPD, which they hypothesised to be part of a compensation pathway39 but could also result from synthetic resistance.
MRTF/SRF and muscle gene expression
MRTF/SRF activity is widely recognised as important in the control of muscle-specific gene expression. However, there is limited information on changes in the MRTFs in response to changing activity or disease. Human studies have found that MRTF expression increases in response to physical training and declines following a period of detraining.21 22 Previous in vitro and animal studies are consistent with a role for these proteins in the regulation of MHCI expression. For example, expression of a dominant negative MRTF-B in differentiating C2C12 cells40 reduces SRF activity and MHCI expression. Similarly, reduced SRF activity is associated with the reduction in MHCI and increased MHCIIa and MHCIIx following hind limb suspension in rats.41
miRNA in muscle disease
In addition to the reduction in miR-1 expression, the pattern of miRNA expression was also different between patients and controls, suggesting a more general change in miRNA expression. The expression of miR-133 and miR-206 were inversely associated with daily physical activity. This change is consistent with the observations of Nielsen et al42 and Keller et al14 who showed that endurance training suppresses the expression of these and other myo-miRs. The fact that we observed no association of miR-1 and miR-499 with physical activity suggests that factors other than daily physical activity are more important in determining their levels in patients with COPD. Similarly, there was a trend towards reduced miR-499 and miR-208, miRNAs that target myostatin, expression of which is increased in weak COPD patients.43
Limitations of the study
The conclusions drawn from this study are limited by the cross-sectional design that only enables us to detect associations between the gene expression and physiological characteristics. Furthermore, the data are correlative and, although they highlight potential mechanisms, they do not prove causality. It should be noted, however, that the highlighted associations have been demonstrated mechanistically through in vitro and in vivo studies.
We have also proposed that patients have reduced SRF activity but could not make direct measurements of SRF activity in our muscle samples. However, from the reduced MRTF and increased MHCI and miR-1 expression, together with altered SRF localization, it seems reasonable to conclude that SRF activity is reduced.
Conclusions
In this study we show that the expression of miRNAs is different in the quadriceps of patients with COPD than in control subjects. In particular, the expression of miR-1 is reduced and is associated with changes in fibre proportion, probably leading to the observed association with muscle mass and exercise performance. Furthermore, the expression of the MRTFs is also suppressed in the patients and again the expression of one of these genes is associated with disease severity. Together with data from studies identifying the MRTFs and SRF as critical regulators of skeletal muscle phenotype, these findings suggest that reduced activity of the SRF pathway contributes to COPD skeletal muscle dysfunction, in part by downregulating miR-1 expression.
Acknowledgments
We are grateful to Dr Jake Bundy for help with the principal components analysis and statistical analysis.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data Supplement - Manuscript file of format pdf
- Data Supplement - Manuscript file of format pdf
- Data Supplement - Manuscript file of format pdf
- Data Supplement - Manuscript file of format pdf
- Data Supplement - Manuscript file of format pdf
Footnotes
MIP and PRK contributed equally to the study.
Funding This work was funded by the BBSRC, Wellcome Trust and the National Institute for Health Research (NIHR) Respiratory Biomedical Unit at the Royal Brompton Hospital and Imperial College. AL is a BBSRC PhD student, SAN received a Wellcome Trust Fellowship, AD received a NIHR Respiratory Biomedical Unit fellowship and WM is a NIHR Clinician Scientist. NSH is a HEFCE Clinical Senior Lecturer. MIP's salary is part funded by the NIHR Respiratory Biomedical Unit at the Royal Brompton Hospital and National Heart & Lung Institute.
Competing interests None.
Patient consent Obtained.
Ethics approval Ethics approval was provided by Royal Brompton and Harefield NHS Trust research ethics committee.
Provenance and peer review Not commissioned; externally peer reviewed.