Article Text

Abstract
Background Exhausting exercise reduces the mitochondrial DNA (mtDNA) content in the skeletal muscle of healthy subjects due to oxidative damage. Since patients with chronic obstructive pulmonary disease (COPD) suffer enhanced oxidative stress during exercise, it was hypothesised that the mtDNA content will be further reduced.
Objective To investigate the effects of exercise above and below the lactate threshold (LT) on the mtDNA content of skeletal muscle of patients with COPD.
Methods Eleven patients with COPD (67±8 years; forced expiratory volume in 1 s (FEV1) 45±8%ref) and 10 healthy controls (66±4 years; FEV1 90±7% ref) cycled 45 min above LT (65% peak oxygen uptake (V′o2peak) and another 7 patients (65±6 years; FEV1 50±4%ref) and 7 controls (56±9 years; FEV1 92±6%ref) cycled 45 min below their LT (50% V′o2peak). Biopsies from the vastus lateralis muscle were obtained before exercise, immediately after and 1 h, 1 day and 1 week later to determine by PCR the mtDNA/nuclear DNA (nDNA) ratio (a marker of mtDNA content) and the expression of the peroxisome proliferator-activated receptor-γcoactivator-1α (PGC-1α) mRNA and the amount of reactive oxygen species produced during exercise was estimated from total V′o2.
Results Skeletal muscle mtDNA/nDNA fell significantly after exercise above the LT both in controls and in patients with COPD, but the changes were greater in those with COPD. These changes correlated with production of reactive oxygen species, increases in manganese superoxide dismutase and PGC-1α mRNA and returned to baseline values 1 week later. This pattern of response was also observed, albeit minimised, in patients exercising below the LT.
Conclusions In patients with COPD, exercise enhances the decrease in mtDNA content of skeletal muscle and the expression of PGC-1α mRNA seen in healthy subjects, probably due to oxidative stress.
- DNA damage, mitochondria biogenesis, oxidative stress, peroxisome proliferator-activated receptor-γ
- coactivator-1α, quadriceps muscle injury, superoxide dismutase
- COPD mechanisms
- exercise
- oxidative stress
- pulmonary rehabilitation
Statistics from Altmetric.com
- DNA damage, mitochondria biogenesis, oxidative stress, peroxisome proliferator-activated receptor-γ
- coactivator-1α, quadriceps muscle injury, superoxide dismutase
- COPD mechanisms
- exercise
- oxidative stress
- pulmonary rehabilitation
Introduction
Exercise limitation is a frequent complaint of patients with chronic obstructive pulmonary disease (COPD) and a major precipitator of their poor health status. Several cellular and molecular alterations that can contribute to exercise limitation have been described in the skeletal muscle of these patients.1–7 Our group has recently identified several mitochondrial abnormalities, including malfunction of the electron transport chain, abnormal kinetics of the mitochondrion permeability transition pore and excessive production of reactive oxygen species (ROS),8 9 that can also contribute to exercise limitation and skeletal muscle dysfunction in COPD.
In healthy subjects, fatiguing exercise causes a reduction in the mitochondrial DNA (mtDNA) content of skeletal muscle due to oxidative damage whereas low-intensity non-fatiguing exercise does not.10 This has not been investigated in patients with COPD but, because mitochondrial ROS production is augmented in such patients,8 11–13 we hypothesised that the fall in mtDNA content reported in healthy subjects10 will be enhanced in patients with COPD, particularly during fatiguing exercise above their lactate threshold (LT). Accordingly, this study sought to compare the dynamic changes induced by two types of exercise protocols (above and below the LT) in mtDNA content of the vastus lateralis muscle in patients with COPD and healthy controls. To get an insight into the potential molecular mechanisms, we also investigated the amount of ROS produced during exercise, the dynamic changes in the activity of manganese superoxide dismutase (MnSOD; EC 1.15.1.1), a major antioxidant enzyme, and of the expression of peroxisome proliferator-activated receptor-γcoactivator-1α messenger RNA (PGC-1α mRNA), a master regulator of mitochondrial biogenesis14 which appears to be abnormal in COPD.6 A better understanding of mitochondrial changes induced by exercise in the skeletal muscle of patients with COPD may be helpful for the design of improved rehabilitation strategies.
Methods
Population
Patients with COPD (diagnosis according to the GOLD guidelines15) had moderate to severe airflow limitation (forced expiratory volume in 1 s (FEV1) <65% reference value), declared to have ceased smoking at least 6 months before enrolling, had not participated in a rehabilitation programme before and had no contraindication for exercise testing. Controls were non-smoking subjects with normal aerobic capacity and resting pulmonary function without any obvious lung, cardiac or other major disease after reviewing their medical information.
Study design
This is a prospective and controlled study. To determine peak oxygen uptake (V′o2peak), all participants performed an incremental (20 W/min) symptom-limited exercise test on a cycle ergometer (ER-900, Jaeger, Hochberg, Germany) with continuous monitoring of oxygen saturation (Nellcor N-180, Pleasanton, California, USA) and breath-by-breath measurements of ventilation and pulmonary gas exchange (Quark-b2 system, Cosmed, Rome, Italy). Those who developed severe hypoxaemia (Sao2 <87%) during the incremental test were excluded from further investigations. Then, 1–2 weeks later, the effects of two constant exercise protocols, above and below the LT, were investigated. The former entailed 3 min resting, 3 min unloaded pedalling and a total of 45 min pedalling (60 rpm) at 65% of V′o2peak whereas the latter entailed pedalling (60 rpm) at 50% of V′o2peak during 45 min. An arterial blood sample was anaerobically drawn immediately after exercise and analysed for blood gases and lactate concentration (Synthesis 1740, Instrumentation Laboratory, Lexington, Massachusetts, USA). Different subjects participated in the two exercise protocols (table 1) because of the high number of muscle biopsies needed (see limitations).
Characteristics of the study population
Patient characterisation
Spirometry was determined in the seated position with a Masterscope-PFT system (VYASIS, Hochberg, Germany); reported values correspond to those measured 15 min after the inhalation of 200 μg salbutamol. Fat-free mass was measured by bioelectrical impedance (Bodystat 1500, Bodystat Ltd, Douglas, Isle of Man, UK).
Muscle biopsies
Biopsies from the vastus lateralis muscle were obtained with an 18G Tru-Cut biopsy needle (Cardinal Health-España, Madrid, Spain) 2–7 days before the exercise test, immediately after it, and 1 h, 24 h and 1 week later. Two patients with COPD and three controls refused to be biopsied 1 week after exercise. The biopsy specimens were immediately frozen and stored at −80°C until analysis.
PCR measurements
To estimate the mDNA content we quantified the mtDNA to nuclear DNA (nDNA) ratio (mtDNA/nDNA). Total DNA was extracted (QIAGEN GmbH, Hilden, Germany) and quantitated spectrophotometrically. The mtDNA content was measured by PCR (ABI Prisma 7700, Perkin-Elmer, California, USA) and corrected by the simultaneous measurement of a single copy nuclear RNAseP gene. Primers used for the analysis of mtDNA were mt806F (5′-CCACGGGAAACAGCAGTGATT-3′) and mt929R (5′-CTATTGACTTGGGTTAAT CGTGTGA-3′). The TaqMan probe was labelled at the 5′ end with the fluorescent dye FAM (5′TGCCAGCCACCGCG-3′). To quantify nDNA we used a commercial kit (PDARs RNAseP; Applied Biosystems, Carlsbad, CA, USA) and a nDNA-specific fluorescent probe labelled internally using the fluorescent dye VIC. PCR conditions were 2 min at 50°C and 10 min at 95°C, followed by 40 cycles of 15 s of denaturation at 95°C and 60 s of annealing/extension at 60°C. Data are expressed as the mean of three measurements.
The amount of PGC-1α mRNA was also quantified by PCR (ABI Prisma 7700, Perkin-Elmer, Foster City, California, USA). The forward and reverse primers employed were 5′-CACTTACAAGCCAAACCAACAACT-3′ and 5′-CAATAGTCTTGTTCTCAAATGGGGA-3′, respectively. Values were expressed relative to the mean data obtained in controls at rest before exercise.
Oxidative stress
The mtDNA oxidation assay used is based on two premises16: (1) oxidative DNA damage blocks the progression of polymerase and causes decreased DNA amplification; and (2) the probability of oxidative lesions is lower in shorter DNA segments. Thus, the ratio between the amplification of a short (8.9 kb) and a long (207 kb) fragment (relative amplification; RA) is a marker that mirrors mtDNA oxidation (ie, the lower RA value the higher mtDNA oxidation16). The two mtDNA fragments amplified were for the 8.9 kb fragment: mt5999F (5-TCTAAGCCTCC TTATTCGAGCCGA-3) and mt14841R (5-TTTCATCATGCGGAGATGTTGG ATGG-3) and the 207 bp fragment: mt4181F (5-ACTTCCTACCACTC ACCCTA-3) and mt4388R (5-TGATAGGTGGCACGGACAAT-3). PCR was performed as described above and the mean resting value of RA in control subjects was considered the reference value (100% RA).
The amount of ROS produced during exercise was estimated assuming that, in healthy subjects, 2% of mitochondrial V′o2 is diverted into the generation of ROS17 and that this is 2.5 times greater in patients with COPD.8
Finally, the activity of MnSOD in skeletal muscle biopsies was measured by the inhibition of nitroblue tetrazolium conversion by MnSOD into a blue tetrazolium salt method (Sigma-Aldrich, St Louis USA), as described by Beauchamp and Fridovich.18
Statistical analysis
The results are presented as mean±SD unless specified otherwise. Comparisons between groups were performed using the paired t test or two-way ANOVA with two factors (group and time), as appropriate. Contrasts at specific time points were made by the unpaired t test. Correlations between variables of interest were explored with the Pearson linear correlation coefficient. A p value <0.05 was considered significant.
Results
The main results are summarised in table 1. Age, body mass index and free fatty mass index were similar between groups. Airflow limitation was moderate to severe in patients with COPD whereas spirometry was normal in controls. Patients with COPD had moderate hypoxaemia. Aerobic capacity (V′o2peak) was normal in controls and moderately reduced in patients with COPD (table 1). Resting MnSOD concentrations were significantly higher in patients with COPD than in controls (table 1). At rest, when all the patients with COPD were compared with all controls, mtDNA/nDNA and PGC-1α values were significantly lower in the former (mtDNA/nDNA: 1096±196 vs 1236±189, p=0.039; PGC-1α: 0.081±0.33 vs 1.01±0.17, p=0.035). However, the differences did not reach statistical significance when analysed separately by groups according to the specific type of constant exercise test to which they had been allocated (table 1).
Exercise above LT
To complete a total of 45 min pedalling at 65% of their V′o2peak, six patients had to stop twice and three patients three times (all because of leg fatigue) whereas three completed the test at once without stopping. Likewise, four controls stopped once (because of leg fatigue) and six did it without stopping. No patient or control subject reached their V′o2peak at any time during the constant work rate exercise challenge. Lactate levels after exercise were similar in both groups (table 1) and indicate that the intensity of the effort was well above the LT (table 1).
mtDNA/nDNA declined after exercise both in controls and patients with COPD but the changes were larger (p=0.02) in patients with COPD (figure 1, upper panel). As a result, immediately after exercise mtDNA/nDNA had decreased by −246±99 in patients with COPD compared with −167±49 in controls (p=0.036), whereas these figures were, respectively, −424±147 vs −277±66 1 h after exercise (p=0.009) and −367±218 vs −194±110 24 h later (p=0.036). One week after exercise (figure 1, upper panel) mtDNA/nDNA had recovered baseline values in both groups (12±91 vs 42±105, p=0.55). By contrast, the concentration of MnSOD tended to increase after exercise in parallel in both groups (figure 2, upper panel), the changes reaching statistical significance 24 h after exercise (p=0.011 and p=0.014 for patients with COPD and control subjects, respectively) and returning to baseline levels 1 week later. During this time course, differences in MnSOD observed at rest between patients and controls groups were basically unchanged throughout (figure 2, upper panel).

Time course of mean±SEM changes in mitochondrial DNA/nuclear DNA (mtDNA/nDNA) in controls (closed circles) and patients with chronic obstructive pulmonary disease (COPD) (open circles) exercising at high intensity (upper panel) and low intensity (lower panel). *p<0.05; **p<0.01; ***p<0.001 isotime COPD vs controls. LT, lactate threshold.
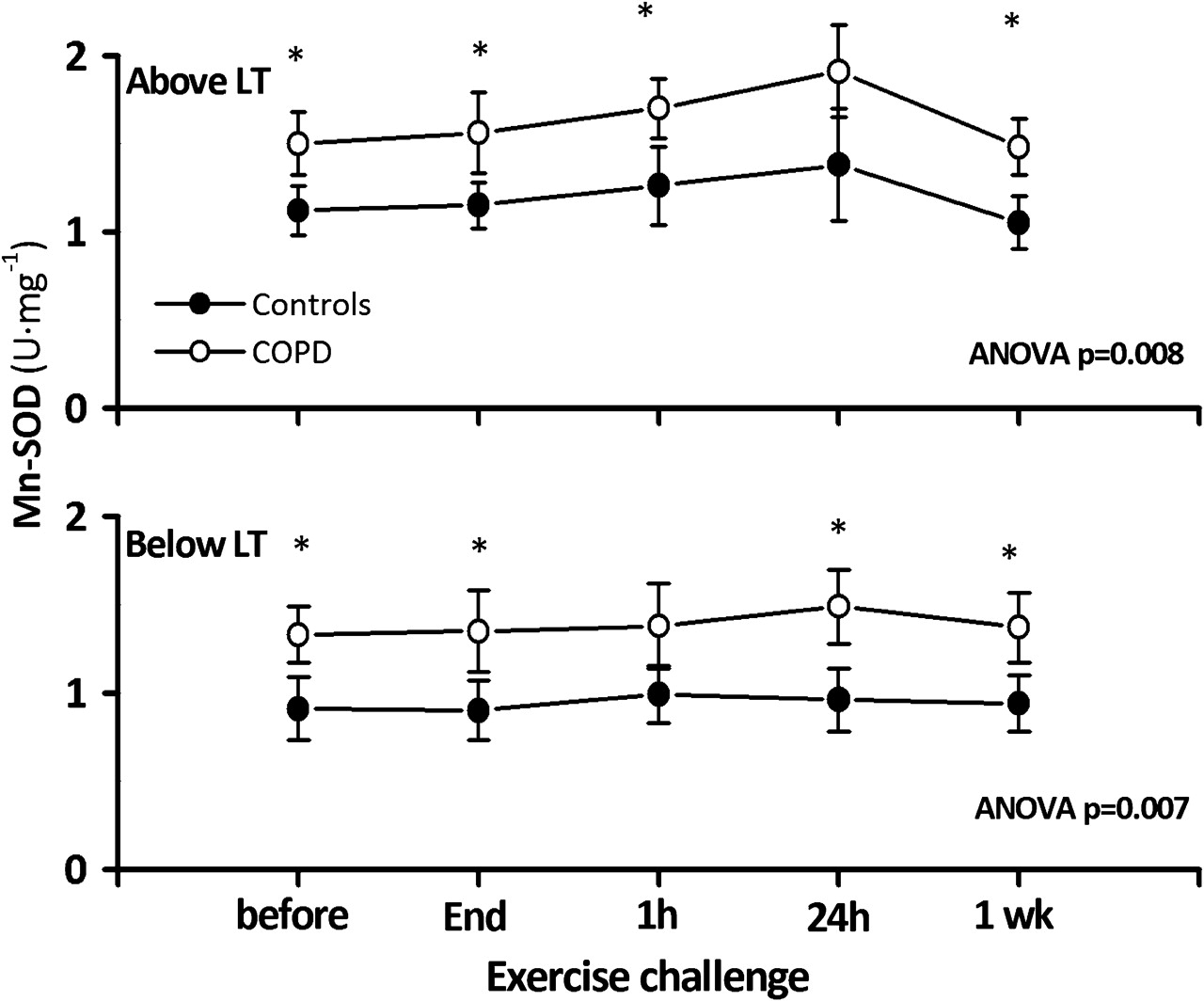
Time course of mean±SEM changes in manganese superoxide dismutase (MnSOD) in controls (closed circles) and patients with chronic obstructive pulmonary disease (COPD) (open circles) exercising at high intensity (upper panel) and low intensity (lower panel). *p<0.05 isotime COPD vs controls. LT, lactate threshold.
PGC-1α mRNA levels at rest (table 1) tended to be reduced in patients with COPD (p=0.07). After exercise (figure 3, upper panel) they increased in both groups to a similar extent (figure 3 upper panel), but the changes were significantly higher (p=0.038) in patients with COPD 1 h after exercise (figure 3, upper panel). By 24 h of recovery, PGC-1α mRNA levels were repressed and corresponded to levels observed at rest (figure 3, upper panel).

Time course of mean±SEM changes in peroxisome proliferator-activated receptor-γcoactivator-1α (PGC-1α) in controls (closed circles) and patients with chronic obstructive pulmonary disease (COPD) (open circles) exercising at high intensity (upper panel) and low intensity (lower panel). *p <0.05; ***p <0.001 isotime COPD vs controls. LT, lactate threshold.
Exercise below LT
The lactate concentration at the end of exercise did not reach 3 mEq/l, confirming that the intensity of exercise was below the LT (table 1). As shown in figure 1 (lower panel), low intensity exercise also induced significant changes in mtDNA/nDNA with respect to baseline in patients with COPD (−109±94 immediately after exercise, p=0.048; −220±91 1 h later, p=0.06; and −159±213 after 24 h, p=0.014), but not in controls (−33±44, p=0.12 immediately after exercise; −73±205, p=0.38 1 h later; and −33±94 after 24 h, p=0.39). It should be noted also that absolute mtDNA/nDNA changes were smaller when patients with COPD exercise below the LT than above it (p=0.007, p=0.024, p=0.004 and p=0.131, respectively, for intergroup comparisons at the end of the exercise and 1 h, 24 h and 1 week later; figure 1). On the other hand, the MnSOD concentration was basically constant after exercise below the LT, so differences with controls were maintained through time (figure 2, lower panel) whereas changes in PGC-1α mRNA were still evident in patients with COPD but absent in controls (figure 3, lower panel).
Potential associations
Because previous studies in healthy subjects ascribed a key pathogenic role to oxidative stress in the observed decrease in the mtDNA content in skeletal muscle,10 we explored potential relationships between the estimated production of ROS during exercise and changes from rest to 1 h after exercise in mtDNA/nDNA in the entire population of subjects (patients and controls) studied both above and below the LT (figures 4 and 5). Our data confirm that a higher production of ROS during exercise was associated with a significant decrease in mRNA/nDNA (figure 4A) and a larger change in MnSOD (figure 4C). Furthermore, we found a good correlation between the estimated ROS production and mtDNA relative amplification (figure 4B), suggesting that the former is a reasonably good proxy of the amount of oxidative stress during exercise.

Correlation between estimated production of reactive oxygen species (ROS) during exercise and changes from rest to 1 h after exercise in (A) mitochondrial DNA/nuclear DNA (mtDNA/nDNA) ratio, (B) mtDNA oxidative damage (relative amplification), (C) manganese superoxide dismutase (MnSOD) and (D) peroxisome proliferator-activated receptor-γcoactivator-1α (PGC-1α) mRNA in controls (closed symbols) and patients with chronic obstructive pulmonary disease (COPD) (open symbols) studied both above (circles) and below the lactate threshold (triangles). FFM, fat-free mass.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlation between measured total oxygen consumed during exercise and changes from rest to 1 h after exercise in (A) mitochondrial DNA/nuclear DNA (mtDNA/nDNA) ratio, (B) mtDNA oxidative damage (relative amplification), (C) manganese superoxide dismutase (MnSOD) and (D) peroxisome proliferator-activated receptor-γcoactivator-1α (PGC-1α) mRNA in controls (closed symbols) and patients with chronic obstructive pulmonary disease (COPD) (open symbols) studied both above (circles) and below the lactate threshold (triangles). FFM, fat-free mass.
On the other hand, we extended previous observations by showing that the estimated ROS production during exercise was also associated with a significant increase in PGC-1α mRNA 1 h after exercise (figure 4D). Likewise, we observed significant correlations between the total amount of oxygen consumed per kg of fat-free mass during exercise and these same muscle variables (figure 5). Finally, we found that changes in PGC-1α mRNA were also significantly related to changes in mtDNA/nDNA (r=0.52, p<0.001) and MnSOD (r=0.40, p=0.017) (data not shown).
Discussion
Our results confirm that, in healthy subjects, fatiguing exercise induces a fall in skeletal muscle mtDNA content10 and upregulates PGC-1α mRNA expression.14 19 We extend these observations for the first time to patients with COPD where, as hypothesised, changes are enhanced (figures 1 and 3), probably in relation to augmented oxidative stress (figures 4 and 5). We also highlight the different recovery dynamics of PGC-1α mRNA expression and mtDNA changes; the former returns to normal values 24 h after exercise (figure 3) whereas the latter takes longer to recover (figure 1). Our results also show that these effects are less marked when patients exercise below the LT (figures 1–3).
Previous studies
A number of cellular and molecular abnormalities have been described in the skeletal muscle of patients with COPD1 4 20 21 including mitochondrial changes.1 3 5 8 9 The lower PGC-1α mRNA expression observed in skeletal muscle biopsies of patients with COPD at rest are in keeping with these previous observations and may be the consequence of inactivity or a switch towards type II fibres (known to express less PGC1α-mRNA and reported in COPD).1 8 20 21 To our knowledge, however, this is the first study that has specifically investigated the effects of exercise, both below and above the LT, on these markers in these patients. Overall, the pattern of response to exercise of mDNA/nDNA and PGC-1α mRNA observed in patients with COPD is similar to that described in healthy subjects10 14 19 but, for a given exercise intensity, mDNA/nDNA changes are magnified in COPD (figures 1 and 3).
Potential mechanisms
At rest the mtDNA/nDNA ratio was lower in patients with COPD. This is consistent with several abnormalities previously described in these patients including decreased density of mitochondria in the vastus lateralis muscle,3 reduced citrate synthase activity4 8 21 22 and/or a higher proportion of type II fibres1 with less oxidative capacity and lower PGC-1α-mRNA expression23 (as was also observed in our patients).
We confirmed that the mtDNA content decreases in healthy subjects after fatiguing exercise10 24 and we show for the first time that this is enhanced in patients with COPD. As previously suggested,10 24 our results support a pathogenic role of oxidative stress because: (1) it is well established that patients with COPD produce a larger amount of ROS than healthy controls during exercise11–13 24; (2) we found both temporal and statistical correlations between the estimated ROS burden produced during exercise and changes in mtDNA content (figure 4); (3) we observed a biological gradient between the expected ROS production and the changes in mtDNA/nDNA that were reduced in patients with COPD exercising below their LT (figure 1) who also have less oxidative stress (table 1 and figures 4 and 5); (4) the fall in mtDNA after exercise was related to changes in MnSOD concentration and PGC-1α mRNA content (figure 4), both known to be related to oxidative stress22 25; and (5) it is biologically plausible that, as the main subcellular site of ROS production is the mitochondria,26 27 the concentration of ROS in the mitochondrial matrix is 5–10-fold higher than in the cytosol or nucleus and the level of oxidised bases in mtDNA is 10–20-fold higher than in nDNA.28 In addition, unlike nDNA, mtDNA lacks protective histones26 and is endowed with relatively low DNA repair activity.26 It is therefore entirely plausible29 that enhanced ROS production during exercise in the skeletal muscle of patients with COPD may contribute to amplifying the decrease in mtDNA in these patients (figures 1 and 4). Together these arguments support a mechanistic role for oxidative stress in the enhanced decrease in mtDNA content in the skeletal muscle of patients with COPD after exercise.
Clinical implications
Our observations may be relevant for rehabilitation programmes in COPD because oxidative mtDNA damage and PGC-1α mRNA expression are prerequisites for adaptation to training.10 19 22 30 In this context it is noteworthy that moderate intensity exercise (below the LT) induced both a significant decrease in mtDNA/nDNA and an increase in PGC-1α mRNA expression in the patients with COPD.
The response of PGC-1α mRNA to exercise indicates that, at least in the patients with COPD studied here who were not characterised by a low body mass index (table 1), the first step for mitochondrial biogenesis and type I fibre differentiation is not altered22 and that, consequently, other mechanisms should be implicated in the pathobiology of muscle atrophy in COPD.1 31 This observation also raises concerns about the potential utility of antioxidant supplementation during rehabilitation as they could dampen ROS signalling and interfere with mitochondrial biogenesis.22
Potential limitations
Some potential limitations of our study deserve discussion. First, the sample size was relatively small and biopsy specimens were not available 1 week after exercise in a few subjects. However, we believe that our results are valid because they were internally consistent within each group. Second, different subjects participated in the two exercise protocols (below and above the LT) because the considerable number of biopsies involved in the study precluded other alternatives. However, the baseline characteristics of the patients and controls in both protocols were similar (table 1) and the biological response of healthy subjects to exercise, both above and below the LT, was similar to that previously described. Third, ROS production during exercise was not measured directly but was estimated from published data.8 17 However, we think that the correlation observed with several independent variables such as mtDNA/nDNA, MnSOD and oxidised mtDNA support our interpretation of the results. Finally, patients with COPD were treated with β2 agonists but these drugs do not appear to influence mitochondrial biogenesis signals, including PGC-1α.32
Conclusions
Our study shows that exercising at the typical intensity and duration of training sessions in pulmonary rehabilitation programmes produces a significant decrease in the mtDNA content of skeletal muscle in patients with COPD and overexpression of PGC-1α mRNA, probably in relation to enhanced oxidative stress. Interestingly, the changes are minimised but not abolished with non-fatiguing exercise below the LT, and this may be relevant for the training effect seen in these patients training below the LT.
Acknowledgments
The authors thank the participants in this study for their willingness to contribute to the advancement of science. They are also grateful to Professor Antonio L Andreu and Professor Helena García-Arumí from the Centre d'Investigació en Bioquímica i Biologia Molecular, University Hospital Vall d'Hebron Barcelona, Spain for their help in the determination of the mtDNA/nDNA. CIBERES is an initiative of the Instituto de Salud Carlos III (Ministerio de Ciencia e Innovación).
References
Footnotes
Funding Supported by Fondo de Investigaciones Sanitarias (PS09/02391).
Competing interests None.
Patient consent All participants gave written informed consent after being made fully aware of the goals and potential risks of the study.
Ethics approval This study was conducted with the approval of the Committee for Ethics in Human Research of Madrid Sanitary Area 1 and all aspects of the study comply with the Declaration of Helsinki.
Provenance and peer review Not commissioned; externally peer reviewed.