Article Text

Abstract
Background and aims Although increasing evidence suggests involvement of the distal airway in all stages of asthma, it is not known whether structural changes (defined as airway remodelling) occur in the distal airways of subjects with mild asthma and those with atopy. The aim of this study was to compare control subjects and those with mild asthma in relation to fibroblast phenotypes and remodelling in central and distal airways.
Methods Distal and central fibroblasts from controls (n=12) and patients with mild asthma (n=11) were cultured and incubated for 24 h with 0.4% serum, or stimulated with transforming growth factor β1 (TGFβ1). [35S]Sulfate-labelled proteoglycans in culture medium were analysed by ion exchange chromatography and sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Proliferation was measured with crystal violet, and exhaled nitric oxide was measured by the fractional nitric oxide technique.
Results Vesican production from distal fibroblasts was significantly elevated in patients with asthma compared with controls (p<0.001), and the percentage collagen-positive area in distal asthma tissue was also enhanced compared with controls (p<0.01). In addition, distal asthma fibroblasts had reduced proliferation capacity compared with those of controls (by 24%; p<0.01). Furthermore, the alveolar nitric oxide concentration was correlated to distal biglycan and perlecan production of subjects with asthma (r=−0.857, p<0.05 and r=−0.750, p<0.05 respectively)
Conclusion It is shown that centrally and distally derived fibroblasts differ in their proteoglycan production and proliferation between central and distal tissue, and in those with asthma compared with controls. It is also demonstrated that remodelling is present in distal lung of subjects with mild asthma. This may be of importance in airway remodelling and asthma progression.
- Asthma
- remodelling
- pathology
- nitric oxide
- sleep apnoea
Statistics from Altmetric.com
Introduction
In the pathogenesis of asthma, the alterations in the central airways are well characterised and involve inflammatory and connective tissue changes with proposed implications for decline in lung function and increased bronchial hyper-responsiveness. However, there are some recently published data to suggest that the distal airways (ie, <2 mm conducting airways and lung parenchyma) may play a role in the pathophysiology of asthma.1–3 Structural pathological changes are collectively called remodelling. In asthma, typical features include thickening of the reticular epithelial basement membrane, subepithelial fibrosis, epithelial shedding and smooth muscle hypertrophy/hyperplasia.4 In this process, the extracellular matrix-producing fibroblast–myofibroblast phenotypes deposit molecules such as collagen and proteoglycans.5–7 Proteoglycans are a family of sulfated molecules consisting of a core protein that is associated with one or several glycosaminoglycan chains. The matrix-associated proteoglycans are mainly the connective tissue-associated versican, the large basement membrane proteoglycan perlecan and the small proteoglycans biglycan and decorin.8 These groups of molecules are of major importance in tissue elasticity and directly affect fluid balance and collagen assembly, and also the activities and storage of cytokines and growth factors such as transforming growth factor β1 (TGFβ1).9 10 Activation of fibroblasts leads to a shift in fibroblast phenotype. This has been studied in other fibrotic disorders such as kidney fibrosis, systemic sclerosis and idiopathic pulmonary fibrosis.11 12 Interestingly, fibroblasts from different airway levels seem to differ in their phenotype. This was shown, for example, in a study in which cultured nasal and bronchial fibroblasts responded differently to TGFβ1, and their matrix production was compared.13 In addition to changes in matrix turnover, upregulation of exhaled nitric oxide (NO) is also increased in asthma, systemic sclerosis and allergic alveolitis, and this parameter is suggested to be a reflection of the increased inflammation.14 Consequently, fractional exhaled nitric oxide (FeNO) is frequently used in the clinic and is believed to be associated with eosinophilic inflammation in the central airways.15 16 The airway epithelium and leucocytes such as macrophages are sources of exhaled NO, but it is likely that other cells such as fibroblasts may also be of importance in increased NO production.
To explore whether fibroblasts from different airway levels in the human lung have adopted distinct disease-related and location-dependent phenotypes, we cultured primary central and distal fibroblasts from subjects with mild asthma and controls without asthma. The control subjects were divided further into two groups based on whether or not they were atopic. Atopic features may precede the onset of asthma, and this is therefore a patient group of interest. In addition, treatment of allergic rhinitis in subjects with asthma may improve co-existing asthma.17
We hypothesised that distal lung fibroblasts are different from centrally derived fibroblasts regarding proteoglycan production, proliferation and morphology. We also wanted to investigate whether differences in fibroblast phenotypes could be observed between those without asthma and those with mild asthma, and whether they could be correlated to inflammatory parameters such as NO.
Materials and methods
Subjects and sampling of tissue
Twenty-nine individuals were enrolled in the study (13 patients with asthma and 16 controls). In six individuals, fibroblast culture was not established and they were therefore excluded from further analysis. Eleven fibroblast cultures were established from patients with asthma with definitive bronchial hyper-responsiveness, defined as PD20 (provocative dose causing a 20% fall in forced expiratory volume in 1 s (FEV1)) <2000 μg methacholine, and they all had mild asthma according to Global Initiative for Asthma (GINA) guidelines.18 All individuals were free from infections and had not used any corticosteroid treatment from 3 months prior to the start of the study. Healthy age-matched non-smoking subjects were used as controls. They all denied having respiratory symptoms, and were all methacholine challenge test negative. Seven of the controls were non-atopic and five were atopic, as defined by confirmed sensitisation by skin prick test to one or more allergens (birch, timothy, mug wort, cat, dog, and/or house dust mite).19 The atopic subjects also had rhinitis. Central bronchoscopy was performed with a flexible bronchoscope (Olympus IT60, Tokyo, Japan) followed by a transbronchial biopsy specimen taken with biopsy forceps (Olympus FB211D) guided by a fluoroscope. As local anaesthesia, lidocaine spray was given through a spray catheter. All subjects gave written informed consent and the study was approved by the ethics committee of Lund University (LU412-03). For more information see the supplementary data online.
Cell cultures and morphological characterisation
For morphological characterisation, crystal violet staining was used as previously described.10 For further characterisation of fibroblasts, cells were labelled with antibodies to vimentin, prolyl-4-hydroxylase or SM22, and incubated with appropriate fluorescent-labelled secondary antibodies. For counting of cellular extensions, 250 cells/subject were randomly counted. For more information see the supplementary data online.
Trichrome staining
Sections of periodate–paraformaldehyde–lysine (PLP)-fixed transbronchial biopsies were stained with Masson's trichrome stain for measurement of collagen content in the alveolar parenchyma. Blue collagen staining was analysed by computerised image analysis (ImageJ, version 1.41, NIH). For a detailed description, see the supplementary data online.
Proliferation assay
Cells were seeded in 96-well plates. Non-adherent cells were washed out after 6 h of incubation and fresh medium was added for the remaining 24 h. Cells were fixed, and stained with crystal violet. For more information see the supplementary data online.
Proteoglycan analysis
Cells were grown in 35S-supplemented medium, and diethyl-aminoethyl (DEAE)-52 cellulose anion exchange resin columns were used to extract the proteoglycans. The individual proteoglycans were separated further on a sodium dodecyl sulfate (SDS)–polyacrylamide gel and quantification of the amount of proteoglycan was done using a scintillation counter as previously described; see the supplementary data online.
NO measurement
Measurements were performed as previously described.18 Briefly, FeNO measurements were done prior to bronchial challenge testing at a flow rate of 50, 100, 200 and 400 ml/s using a NIOX nitric oxide analyser (Aerocrine AB, Stockholm, Sweden), and the results were expressed as parts per billion (ppb). Alveolar NO concentration and bronchial flux of NO were calculated with a two-compartment linear model using a flow rate of 100–400 ml/s.
Statistical methods
For non-parametric data, between-groups comparisons were performed with the Kruskal–Wallis test (a non-parametric test on ranks, which compares the means of three or more unmatched groups) with the Dunn multiple comparison test followed by the Mann–Whitney U test. Correlations were tested by Spearman rank correlation using Graph pad software version 5.03 (San Diego, California, USA). For parametric data the Student t test was used. If not all subjects are represented in the graphs, these have been excluded due to technical problems such as loss of biopsies when staining tissue or limited amount of material. Data are presented as median values together with the range unless otherwise stated; p values of <0.05 (*) and <0.01 (**) denote significant levels of difference.
Results
Patient characterisation and fibroblast recovery
Of the 29 individuals originally considered for the study, fibroblast cultures were established from 23 (11 patients with asthma and 12 controls). Subjects with asthma were 22–38 years of age (mean 28) and controls were 22–40 (mean 28). No differences in FEV1 or FEV1% predicted were seen when comparing those with asthma and controls (4.2 vs 3.9 litres and 95% vs 104%, respectively). However, the mean FeNO50 (exhaled NO at 50 ml/s) of those with asthma was significantly elevated relative to that of the controls (75.8 (15.8–240) ppb vs 15.4 (8.4–27.7) ppb), p=0.017) and relative to that of the atopic controls without asthma (75.8 (15.8–240)) ppb vs 16.6 (9.8–27.7) ppb), p=0.018). For bronchial flux of NO, subjects with asthma had levels that were sixfold higher than those of the controls (4.61 (0.68–12.4) vs 0.73 (0.38–1.28), p=0.007) and also higher compared with atopic controls (4.61 (0.68–12.4) vs (0.81 (0.47–1.28), p = 0.024 (table 1).
Clinical characterisation of subjects included in the study and fibroblasts successfully cultured from different locations of the lung from these subjects
Increased remodelling in tissue from distal airways in patients with mild asthma
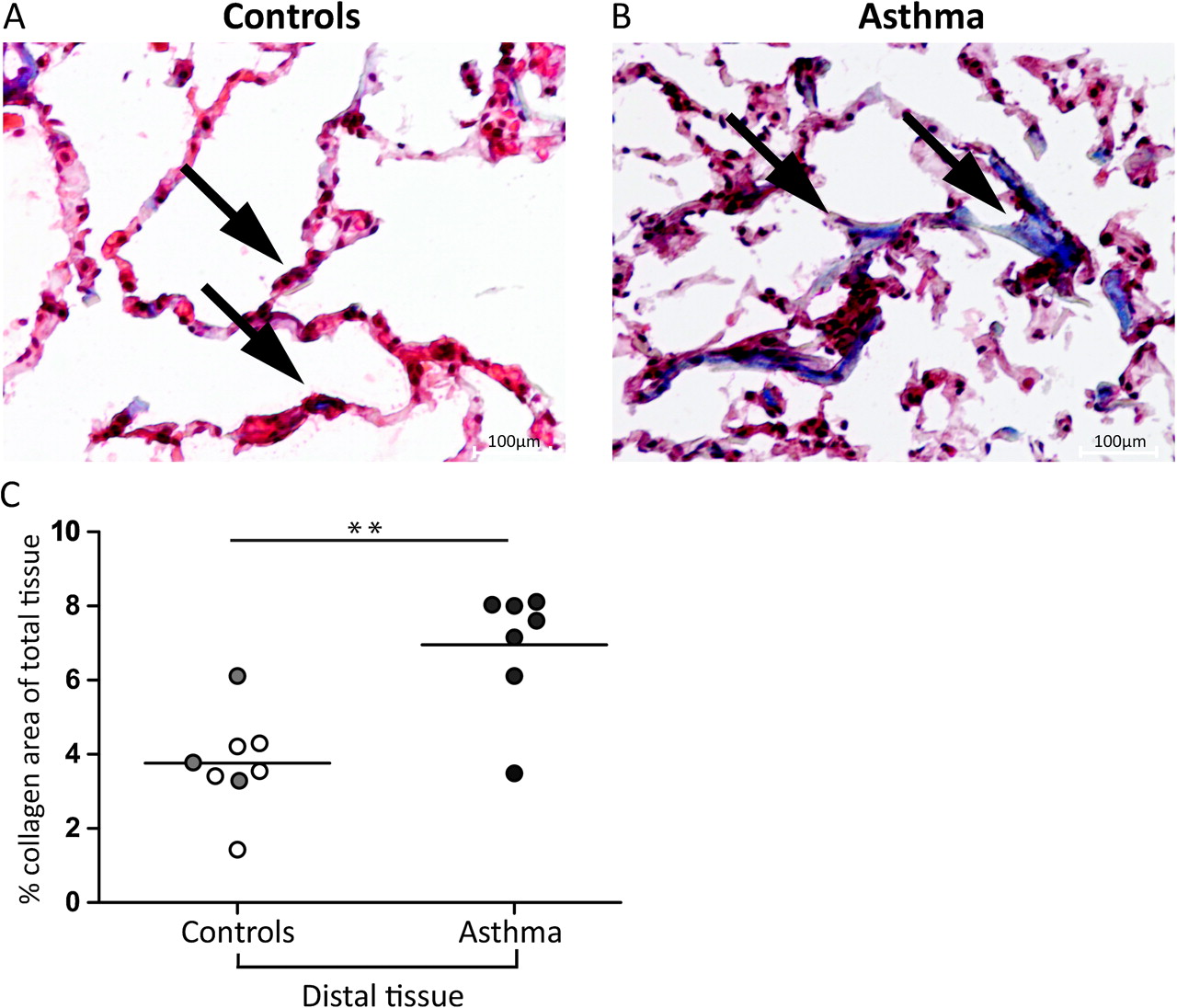
Distal lung biopsies were stained by trichrome staining to visualise collagen regions in the alveolar parenchyma (exemplified in figure 1A,B, black arrows). Elongated spindle-shaped collagen areas were clearly visualised in distal biopsies especially from patients with asthma (figure 1B). Distal lung collagen expression was significantly elevated in patients with asthma compared with controls (6.9 (3.6–8.1) vs 3.8 (1.42–6.2), p=0.007). No difference was seen between atopic and non-atopic controls (figure 1C).

Collagen in distal airways in patients with mild asthma. Representative images of collagen in distally derived biopsies from a control subject (A) and a patient with asthma (B) visualised with Masson trichrome staining ×400 magnification. Black arrows indicate collagen (blue) in the parenchyma from a control and a patient with asthma. Quantitative data of tissue collagen content in controls and subjects with asthma are shown in (C). Mann–Whitney tests were used to calculate p values. Values are expressed as collagen-positive area as a percentage of the total tissue area. Because of loss of biopsies (detached from the slide) in the staining procedure collagen was not analysed in two of the atopic subjects and in two of the non-atopic controls. Open circles represent non-atopic controls (n=5), grey circles represent atopic controls (n=3) and filled circles represent subjects with asthma (n=7). Mann–Whitney tests were used to calculate significant differences between controls and patients with asthma.
Characterisation of fibroblasts
Distal fibroblasts were all positive for the filament proteins vimentin (figure 2A,E) and α-smooth muscle actin (figure 2B,F). Furthermore, these cells also expressed prolyl-4-hydroxylase, which is a key enzyme required for the post-translational hydroxylation of proline residues in collagen (figure 2C,G). The cells were negative for one of the most sensitive markers of smooth muscle cells, SM22, further supporting the idea that we had cultured a homogenous fibroblast population (figure 2D,H). The same pattern of staining was seen for the centrally derived fibroblasts (data not shown), which has also been shown previously.35

Characterisation of distal fibroblasts. Distal fibroblasts were cultured for 48 h and stained for mesenchymal and smooth muscle proteins to investigate fibroblasts from controls and patients with asthma. Cells from control individuals (A–D) and patients with asthma (E–H) were all positive for the mesenchymal fibroblast protein vimentin (green, Alexa 488), α-smooth muscle actin (yellow, Cy3) and prolyl-4-hydroxylase (red, Alexa 647) but negative for the smooth muscle marker SM22 (green, Alexa 488). Staining of nuclei is visualised in blue (4',6-diamidino-2-phenylindole (DAPI)).
Distally and centrally derived fibroblasts from subjects with asthma have reduced proliferation capacity
Centrally derived fibroblasts from subjects with asthma had 24% lower proliferation capacity relative to the controls, after 24 h (1.06 (0.96–1.15) vs 1.39 (1.08–1.88), p=0.004) (figure 3A).This was also seen with distally derived asthma fibroblasts, which had a lower proliferation rate (23%) compared with the corresponding controls (1.27 (1.12–1.57) vs 1.65 (1.23–2.06), p=0.002) (figure 3A). There was a significant increase in fibroblast proliferation when comparing central versus distal in both controls and subjects with asthma (1.39 (1.08–1.88) vs 1.65 (1.23–2.06), p=0.034, and 1.06 (0.96–1.15) vs 1.27 (1.12–1.57), p=0.004, respectively) (figure 3A). The same difference in proliferation could also be measured after 48 h (data not shown).

Culture and morphological characterisation of centrally and distally derived fibroblasts. (A) Fibroblasts from central and distal locations were seeded in 96-well plates at a density of 5000 cells/well and cultured for 24 h. Glutaraldehyde-fixed cells stained with crystal violet were lysed with 1% Triton X-100 and the absorbance measured at 595 nm. (B) Each cell was counted for cellular extensions (black arrows). In two of the patients with asthma, proliferation of centrally derived fibroblasts could not be measured because of a limited amount of material. Student t test was used to calculate p values. Open circles represent non-atopic controls, grey circles represent atopic controls and filled circles represent subjects witha sthma. *, Significant difference (p<0.05) when comparing proliferation of centrally derived versus distally derived fibroblasts from controls. **, Significant difference (p<0.01) when comparing distally derived fibroblasts from controls versus patients with asthma, when comparing centrally control fibroblast versus centrally asthma fibroblasts, and when comparing central asthma versus distal asthma fibroblasts.
Central fibroblasts from subjects with asthma have more cellular extensions than distal fibroblasts
When counting the number of cellular extensions in centrally derived fibroblasts, cells from subjects with asthma had significantly more (1.4 times more) cellular extensions than those from controls (8.3 (4–20) vs 5.9 (2–20), p<0.001), figure 3B, black arrows). Distally derived fibroblasts were more elongated and had fewer extensions than centrally derived fibroblasts (2.6 (1–7) vs 6.6 (2–20), p<0.001).
Increased matrix production by distally and centrally derived fibroblasts is associated with disease
Production of the large molecule versican was significantly greater in centrally derived fibroblasts from those with asthma than in control fibroblasts (354 (201–571) vs 96 (18–201) dpm (disintegrations/min)/μg, p<0.01, figure 4A). Consistent with data from the central airways, distally derived fibroblasts from subjects with asthma also had considerably higher versican production than control fibroblasts (210 (163–259) vs 106 (45–162) dpm/μg, p<0.001, figure 4A). In control subjects, there was no difference between atopic and non-atopic subjects (figure 4A). Expression of the small leucine-rich proteoglycan biglycan was studied further. We found that there was enhanced biglycan production from centrally derived fibroblasts from those with asthma compared with controls (163 (84–346) vs 81 (47–144) dpm/μg, p=0.006, figure 4B). These differences could not be detected in distal airway fibroblasts. However, biglycan production by distally derived fibroblasts from patients with asthma was significantly lower than from non-atopic controls (145 (72–207) vs 349 (224–480) dpm/μg, p<0.001, figure 4B). Interestingly, atopic controls also had significantly lower production of biglycans than non-atopic controls (122 (40–255) vs 349 (224–480) dpm/μg, p=0.010) (figure 4B). The large (>400 kDa) multidomain proteoglycan perlecan which is mainly localised in the basement membrane was not different in expression regardless of whether central or distal localisation or different groups were compared (figure 4C). As observed for distal biglycan production, decorin production in both patients with asthma and atopic controls was significantly lower than in non-atopic controls (127 (56–258) and 109 (66–154) vs 343 (55–476) dpm/μg; p=0.026 and p=0.048, respectively) (figure 4D).

Proteoglycan production in centrally and distally derived fibroblasts. Centrally and distally derived fibroblasts from controls and from subjects with asthma were analysed for proteoglycan production: (A) versican; (B) biglycan; (C) perlecan; and (D) decorin. Cells were grown in 35S-supplemented sulfate-poor medium, and diethyl-aminoethyl (DEAE)-52 columns were used to extract the proteoglycans. Quantification of proteoglycan production was done using a scintillation counter and is presented as dpm/μg protein. The individual proteoglycans were separated further by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE). Mann–Whitney tests were used to calculate p values.Values presented are medians, and open circles represent non-atopic controls (central n=3, distal n=7), grey circles represent atopic controls (central n=4, distal n=5) and filled circles represent subjects with asthma (central n=8, distal n=7). **, Statistically significant difference (p<0.01) for versican (A) and biglycan (B) production by centrally derived fibroblasts when comparing controls and patients with asthma. ***, Significant difference (p<0.001) for distal versican production (controls vs asthma patients). * and #, Significant difference (p<0.05) for central biglycan production versus distal production from controls and for distal biglycan and decorin production (non-atopic controls vs patients with asthma (open circle vs filled circle) and non-atopic controls versus atopic controls (open circle vs grey circle)).
Alveolar NO was correlated to production of individual proteoglycans in distal airways
Negative correlations were observed when comparing alveolar NO concentrations and distal biglycan and perlecan production by fibroblasts from patients with asthma (r=−0.857, p=0.033, and r=−0.750, p=0.012, respectively) (figure 5A,B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Biglycan and perlecan production by distally derived fibroblasts from subjects with asthma was plotted against alveolar nitric oxide (NO) concentration (A and B, respectively) in subjects with asthma (filled circle). Spearman correlation was used to calculate the significance of the degree of association between proteoglycan production and alveolar NO concentration.
Asthma fibroblasts respond less to TGFβ1 stimulation than those from controls
TGFβ1 significantly increased the production of the proteoglycans versican, biglycan and perlecan in both centrally and distally derived control fibroblasts compared with unstimulated cells (centrally derived (p=0.016, p=0.016 and p=0.039, and distally derived (p=0.039, p<0.001 and p=0.034), respectively) (table 2). In centrally derived asthma fibroblasts, versican production was increased compared with unstimulated cells (p=0.039). In distal airways, TGFβ1 stimulation significantly increased biglycan production from asthma fibroblasts (p=0.031). For individual values after TGFβ1 stimulation, see the supplementary files.
Influence of transforming growth factor β1 (TGFβ1) on individual proteoglycan production from centrally and distally derived fibroblasts and related to basal level production (0.4% serum)
Discussion
In this study we have demonstrated that both distally and centrally derived fibroblasts from patients with asthma have an altered cell proliferation and production of matrix molecules, such as versican, compared with controls. We also verified distal airway remodelling in subjects with mild asthma, as described by increased collagen accumulation in peripheral airway tissue. We further showed that distally and centrally derived fibroblasts have different phenotypes, an observation that strengthens the hypothesis that the nature of distal airway remodelling differs from brochial remodelling. Moreover, our results also show that production of the proteoglycans perlecan and biglycan can be linked to alveolar exhaled NO levels.
Remodelling in distal lung
It is well established that airway remodelling and inflammation take place in the subepithelial compartment in central airways of subject with atopy and asthma.4 20 21 It is also known that inflammation occurs in the distal airways and lung parenchyma.22 However, this is, to the best of our knowledge, the first study demonstrating remodelling in the distal airways in subjects with mild asthma, as shown by increased deposition of collagen. In this context, the differences in versican production by distal airway fibroblasts in our in vitro experiments further strengthen the hypothesis that fibroblasts also play a role in distal asthma pathophysiology. The present demonstration of remodelling in the distal lung already in mild asthma may be an important observation that possibly has implications for disease progression since remodelling is a feature in both moderate and severe asthma. Higher levels of versican production were only seen in individuals with asthma, suggesting a shift in disease-related fibroblast phenotypes. It has been proposed that due to its high chondroitin sulfate/dermatan sulfate content, versican is destructive for the elasticity in the lung by inhibiting elastin-binding protein (EBP), and thus interferes with assembly of the elastic fibres.23 Accumulation of versican in those with mild asthma could consequently affect lung stiffness in both the central and the distal airways, and thus directly influence lung function.5 The negative correlation of alveolar NO concentration and the proteoglycans biglycan and perlecan suggests that NO may also reflect the degree of remodelling. A lower perlecan production might directly influence basement membrane integrity. Furthermore, the ability of perlecan to bind cytokines and affect epithelial adherence and function may also play an important role in distal remodelling processes.24
Fibroblast phenotypes in distal airways—a link to exhaled NO?
In this study, there was a correlation between the levels of exhaled NO and proteoglycan production in distal airways. This finding links remodelling to clinical parameters and suggests that NO may be used as a marker for fibroblast activity. NO, which is a mediator molecule involved in both proinflammatory and anti-inflammatory processes, is known to be upregulated in asthma.10 Although the production of NO in patients with asthma is believed to be due to expression of inducible nitric oxide synthase (iNOS) in epithelial cells, the primary source has not been identified yet. NO has previously been suggested to be involved in lung fibrosis and fibroblast differentiation.25 26 It is also known that products in the NO cycle can digest glycosaminoglycans into biologically active fragments. This process could possibly be controlled by fibroblasts due to their expression of iNOS, and thus be responsible for the increased NO production in patients.27 28 Elevated levels of NO have been shown in patients with more severe asthma, and these patients are known to have more evidence of remodelling in the distal airways.29 Moreover, alveolar NO has also been shown to be elevated in those with mild asthma as compared with controls without asthma.19
Role of small proteoglycans in airway remodelling
In this work basal biglycan production was also altered in centrally derived asthma fibroblasts, further supporting the hypothesis that the composition of connective tissue may be changed in mild asthma. Notably, both biglycan and decorin production from distally derived fibroblasts from atopic individuals with rhinitis was significantly lower than in non-atopic controls, which indicates that these individuals have changes in distal connective tissue composition comparable with those in subjects with asthma. This suggests involvement of the upper and distal airways in airway diseases such as seasonal and chronic rhinitis. However, these observations require confirmation in studies with adequate patient numbers for the analyses to be robust. The important role of proteoglycans in asthma, fibrosis, and chronic obstructive pulmonary disease (COPD) and other diseases is well documented, at both the cellular and the physiological level.2 23 30 Previously published data have shown that the small proteoglycans biglycan and decorin induce morphological and cytoskeletal changes in fibroblasts, suggesting that they have a role in conversion of the resident fibroblast to an inflammatory fibroblast phenotype promoting wound healing processes.10 This role of biglycan has also been discussed by others in the context of biglycan functioning as a proinflammatory molecule involved in innate immunity.31 Expression of proteoglycans in centrally derived tissue sections from atopic subjects and those with fatal asthma and in animal models has previously been studied using immunohistochemistry, further confirming our results.6 7 32 Fibroblast production of collagen, which is the most abundantly expressed extracellular matrix molecule in the lung, was not measured in the present study. However, some of the proteoglycans interact individually with collagen to modulate collagen organisation and network formation.33 One of the most potent mediators in stimulating collagen and proteoglycan production is TGFβ1, and its role in human asthma has been extensively reviewed, but there are contradictory results concerning its direct role. Current evidence suggests that expression of TGFβ1 is upregulated in asthma.34 Overall, since no differences in increased proteoglycan production were observed between groups, our results suggests that the fibroblast changes with regard to proteoglycan production are already present in the cells at basal levels, and might only be enhanced by TGFβ1.
In this study we also observed a shift from a proliferating cell phenotype, as seen in centrally derived fibroblasts from controls, to a resident matrix-producing cell in subjects with asthma, with increased numbers of cellular extensions. This indicates the presence of different fibroblast phenotypes in controls and patients with asthma. It is an interesting observation that might suggest that fibroblasts in asthma acquire a more activated contractile phenotype, although this needs to be investigated further.35 Furthermore, our data are in contrast to data published by Wenzel and colleagues where no difference was observed between those with asthma and controls except for localisation.1 This could be due to a different experimental set-up as they used postmortem lung material as the source of control tissue.
Summary
In conclusion, we have shown for the first time that matrix production by both centrally and distally derived fibroblasts is different for those with mild untreated asthma and for controls. In addition, remodelling in distal lung is also present in subjects with mild asthma as visualised by increased collagen. These differences could also be linked to exhaled NO, which suggests an association between connective tissue composition and lung function in those with mild asthma. In the delicate balance of physiologically normal responses to inflammation and the resolving processes that follow, proteoglycans may have a pivotal role in the initiation and maintenance of pathological remodelling in the airways. Further studies in patients with other subtypes of asthma and comparative studies in other lung diseases will be required to define the precise mechanisms of matrix involvement in order to intervene and counteract this process in the human lung.
Acknowledgments
The authors thank Lena Thiman for her laboratory skills and technical assistance.
References
Supplementary materials
Web Only Data thx.2009.129320
Files in this Data Supplement:
Web Only Data thx.2009.129320
Files in this Data Supplement:
Footnotes
Funding This work was supported by grants from the Swedish Research Council (11550, 7970), the Heart and Lung Foundation, Centrala Försöksdjursnämnden (CFN), the Greta and John Kock Foundation, the Alfred Österlund Foundation, the Crafoord Foundation, Riksföreningen mot Rheumatism, the Schyberg Foundation, the Hedberg Foundation, the Gustaf V 80th Fund, Evy and Gunnar Sandberg Foundation and the Medical Faculty of Lund University.
Competing interests None.
Patient consent Obtained.
Ethics approval This study was conducted with the approval of the the ethics committee of Lund University (LU412-03).
Provenance and peer review Not commissioned; externally peer reviewed.