Article Text

Abstract
Background 15-S-Hydroxyeicosatetraenoic acid (15(S)-HETE) and 13-S-hydroxyoctadecadienoic acid (13(S)-HODE), both of which are metabolites of 15-lipoxygenases (15-LOXs), are endogenous ligands for peroxisome proliferator-activated receptor gamma (PPARγ). The activation of PPARγ inhibits cell growth and induces apoptosis in some cancers. The role of 15(S)-HETE) and 13(S)-HODE in the development of lung cancer is not clear.
Methods 15-LOXs, 15(S)-HETE and 13(S)-HODE were monitored during the development of mouse lung tumours induced by the tobacco smoke carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) and the levels of these markers were determined in 54 human non-small cell lung cancers.
Results 15-LOXs, 15(S)-HETE and 13(S)-HODE levels were significantly reduced in human lung cancer tissue compared with non-tumour lung tissue (p=0.011 and p=0.022, respectively). In mouse experiments, 15(S)-HETE and 13(S)-HODE started to reduce at 26 and 30 weeks, respectively, after NNK treatment. The time frame of 15(S)-HETE reduction was in line with the decrease in 12/15-LOX mRNA and protein. A significant difference in the number of tumours in NNK-treated mice and controls was not observed until week 34 (p<0.05) and week 38 (p<0.01). The reduction in 12/15-LOX and 15(S)-HETE therefore predated the appearance of lung tumour. Furthermore, PPARγ activity was decreased in NNK-treated mouse lungs compared with the control, and the decreased PPARγ activity occurred at the same time points as the reduction in 12/15-LOX and 15(S)-HETE.
Conclusion These findings indicate that the reduction in 15-LOX, 15(S)-HETE and 13(S)-HODE results in the decreased PPARγ activity seen in lung tumours and contributes to the development of lung tumours induced by tobacco smoking.
- Lung tumour
- 15-lipoxygenases
- 15(S)-HETE
- 13(S)-HODE
- PPARγ
- lung cancer
- non-small cell lung cancer
Statistics from Altmetric.com
Introduction
Lung cancer is the most common cause of death from cancer in the world, with an overall 5-year survival rate of only 14% following diagnosis.1 2 Approximately 90% of all lung cancers are directly attributable to smoking. Lung cancer is divided into two major classes based on its biology, treatment and prognosis: non-small cell lung cancer (NSCLC) and small cell lung cancer (SCLC). NSCLC accounts for 80–85% of all cases of lung cancer and it includes three major types: squamous cell carcinoma, adenocarcinoma and large cell carcinoma. Despite advances in early detection and standard treatment, NSCLC is often diagnosed at an advanced stage. The heavy burden of lung cancer and the failure of effective approaches to reduce mortality have resulted in an interest in identifying signature biomarkers for lung cancer and target molecules for antitumour treatment.
Peroxisome proliferator-activated receptors (PPARs) belong to the nuclear receptor family consisting of a group of approximately 50 transcription factors implicated in many different biological processes, and are considered as important targets in the development of new drugs. The binding of ligands to receptors results in change of expression of mRNAs encoded by PPAR target genes. Activation of PPARγ by its endogenous and exogenous ligands can inhibit cell proliferation and growth, and induce differentiation and apoptosis.3–5 Several studies have shown that the activation of PPARγ can inhibit the growth of lung cancer cells.3 5 6 PPARγ protein appears to correlate with the maturational stage of cells with a well-differentiated phenotype in NSCLC.7 Well-differentiated NSCLC tissues frequently express more PPARγ. However, the activation of PPARγ is thought to be defective,3 6–8 despite the high expression of PPARγ in lung cancers.
15-S-Hydroxyeicosatetraenoic acid (15(S)-HETE) and 13-S-hydroxyoctadecadienoic acid (13(S)-HODE), the two main metabolites of 15-lipoxygenases (15-LOXs), are endogenous ligands of PPARγ. They can inhibit cell proliferation and induce apoptosis in several types of human cancer.8–11 Two different types of human 15-LOXs have been found: 15-LOX-1 mainly metabolises linoleic acid (LA) to 13(S)-HODE but also converts arachidonic acid (AA) to 15(S)-HETE; 15-LOX-2 preferentially converts AA to 15(S)-HETE and metabolises LA poorly. Lungs express both of these two 15-LOXs, which suggests that 15-LOXs or their products (15(S)-HETE, 13(S)-HODE) may have a role in maintenance of the differentiated phenotype of lung epithelial cells, and that their abnormal expression levels or functions may contribute to the development and progression of lung cancer.
To our knowledge, 13(S)-HODE and 15(S)-HETE have not been investigated in NSCLC. In the present study we measured the levels of 15-LOX-1 and -2 and their enzymatic activities in human NSCLC. To further study the pathological properties of these two endogenous ligands and PPARγ during lung carcinogenesis, the levels of 15(S)-HETE and 13(S)-HODE and their related enzyme 12/15-LOX (murine orthologue to human 15-LOX-1) were monitored during the course of lung tumour development in A/J mice induced by the tobacco smoke carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK). We wished to determine whether 15(S)-HETE and 13(S)-HODE or their enzymes 15-LOX-1 and -2 contribute to the development of lung tumours, whether they can serve as potential biomarkers in the early detection of NSCLC and whether these two ligands can be used as potent therapeutic targets for NSCLC.
Methods
Materials
ELISA kits for 15(S)-HETE or 13(S)-HODE were purchased from Assay Designs (Ann Arbor, Michigan, USA); RNA extraction kits were supplied by Qiagen (Valencia, California, USA); RT kits were from Promega (Madison, Wisconsin, USA); Sybr Green qPCR SuperMix reagent was from Invitrogen (Carlsbad, California, USA); NNK was obtained from Chemsyn Science Laboratories (Lenexa, Kansas, USA); 12/15-LOX antibodies were from Cayman Chemical (Ann Arbor, Michigan, USA); and antibodies for PPARγ and β-tubulin were from Santa Cruz Biotechnology (Santa Cruz, California, USA).
Patients and tissue samples
NSCLC and their corresponding non-tumour tissue samples were collected from 54 patients, none of whom had previously been treated with chemotherapy or radiation therapy. These tissues were stored in liquid nitrogen until required.
Establishment of lung tumour mouse model
Five-week-old female A/J mice were randomly assigned to two groups for treatment with either phosphate buffered saline (PBS) (controls) or NNK. NNK was dissolved in sterile PBS. Aqueous NNK solution was prepared weekly and stored at 4°C.12 At 6 weeks of age the mice were treated with a single intraperitoneal injection of 100 mg/kg NNK. The animals were killed at 4-week intervals from 26 to 38 weeks after the NNK or PBS injection. Lung tumours induced by NNK were pathologically confirmed to be NSCLC.
Reverse transcription reaction/quantitative real-time PCR (real-time qRT-PCR)
cDNA was generated with 1 μg isolated RNA and real-time qRT-PCR was performed using Sybr Green qPCR SuperMix. The targeted regions of the 15-LOX-1 (295 bp), 15-LOX-2 (170 bp), 12/15-LOX (201 bp) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (136 bp for human, 435 bp for mouse) genes were amplified and cloned into pGEM-T easy vector (Promega). Standard curves were constructed by serial dilution of purified ScaI-linearised plasmids. The difference in the gene expression was calculated according to the standard curve. A non-reverse transcriptase control was set up for each reaction. The expression of genes was normalised using the housekeeping gene GAPDH. The following primer sequences were used: 15-LOX-1, (forward) 5′-GCCCAGGACCGAGGGTTTC -3′ and (reverse) 5′-GGCCCACAGCCACCATAAC-3′; 15-LOX-2, (forward) 5′-TTGCAGGCTGAGCTAGAGAAGG-3′ and (reverse) 5′-CTG AGCTGGATGGCGAGAGG-3′; 12/15-LOX (forward) 5′-ATCGGTACGTGGGAATG-3′, 12/15-LOX (reverse) 5′-GCGCAGTGCATGTGAAGATG-3′; human GAPDH (forward) 5′-GGGTGTGAACCATGAGAA-3′ and (reverse) 5′-GACTGTGGTCATGAGTCCT-3′; and mouse GAPDH (forward) 5′-CACTGCCACCCAGAAGACTG-3′, GAPDH (reverse) 5′-TCCACCACCCTGTTGCTGTA-3′. Real-time PCR reactions consisted of the following steps: 50°C 2 min, 95°C 10 min, 38 cycles at 95°C 15 s and 60°C 60 s.
Quantitative determination of 15(S)-HETE and 13(S)-HODE
15(S)-HETE and 13(S)-HODE were quantitatively measured using enzyme immunoassay kits from Assay Designs and the assays were performed according to the manufacturers' instructions. Concentrations of 15(S)-HETE and 13(S)-HODE were expressed in pg/mg wet weight of lung tissue.
Histological examination
Mice were killed every 4 weeks starting at week 26. The lungs of all mice were collected and underwent gross inspection. The left lung was inflated with 10% formalin and the whole lung was fixed. In order to examine the different parts of the lung, it was cut into four blocks to obtain tissue from different positions. Four 5 μm paraffin sections were then generated from these four tissue blocks for staining with haematoxylin and eosin (H&E).
Western blot analysis
Approximately one-third of frozen right lung tissue from A/J mice was pulverised in a mortar containing liquid nitrogen. The protein was isolated and western blot analysis was performed as described previously.13 14
Immunohistochemical analysis
Immunohistochemical staining was performed according to the standard procedure.15 Negative controls were prepared by using normal rabbit serum instead of the primary antibody. The intensity of the antigen staining was scored according to the standard described in table 1.
Scoring standard for antigen staining
PPARγ transcriptional activity
The transcriptional activity of PPARγ was measured using a kit from Cayman Chemical and the assay was performed according to the manufacturer's instructions. This kit is specific to PPARγ and does not cross-react with PPARδ or PPARα.
Statistical analysis
Paired t tests were performed to compare the difference in 15(S)-HETE, 13(S)-HODE and 15-LOX-1 and -2 concentrations between non-tumour and tumour lung tissues. The results of H&E staining and immunohistochemical staining of mouse lung tissue were analysed using a non-parametric test (Mann–Whitney) and the results of ELISA, real-time qRT-PCR and PPARγ transcriptional activity were evaluated by a parametric test (Student t test). Parametcir data confirmed the normality distribution before the statistical procedures were performed and are presented as mean±SE. Semi-quantified data of 12/15-LOX immunohistochemistry staining are presented as median values. All tests were two-sided and p values <0.05 were considered to be significant. Asterisks shown in the figures indicate significant differences between experimental groups and the corresponding controls (*p<0.05; **p<0.01).
Results
Clinical and pathological information of patients
All 54 lung carcinomas were histologically confirmed to be NSCLC; the clinical and pathological data of the patients are shown in table 2.
Clinical and pathological information of patients (n=54)
Levels of 15(S)-HETE and 13(S)-HODE and their enzyme 15-LOXs in human NSCLC
To determine the levels of 15(S)-HETE and 13(S)-HODE in human NSCLC, we collected 54 human NSCLC and their matched non-tumour tissues to detect levels of these two ligands. 15(S)-HETE and 13(S)-HODE levels were normalised to the wet weight of lung tissues. As shown in figure 1A, the levels of 15(S)-HETE and 13(S)-HODE were significantly higher in the non-tumour lung tissue than in the tumour tissue.
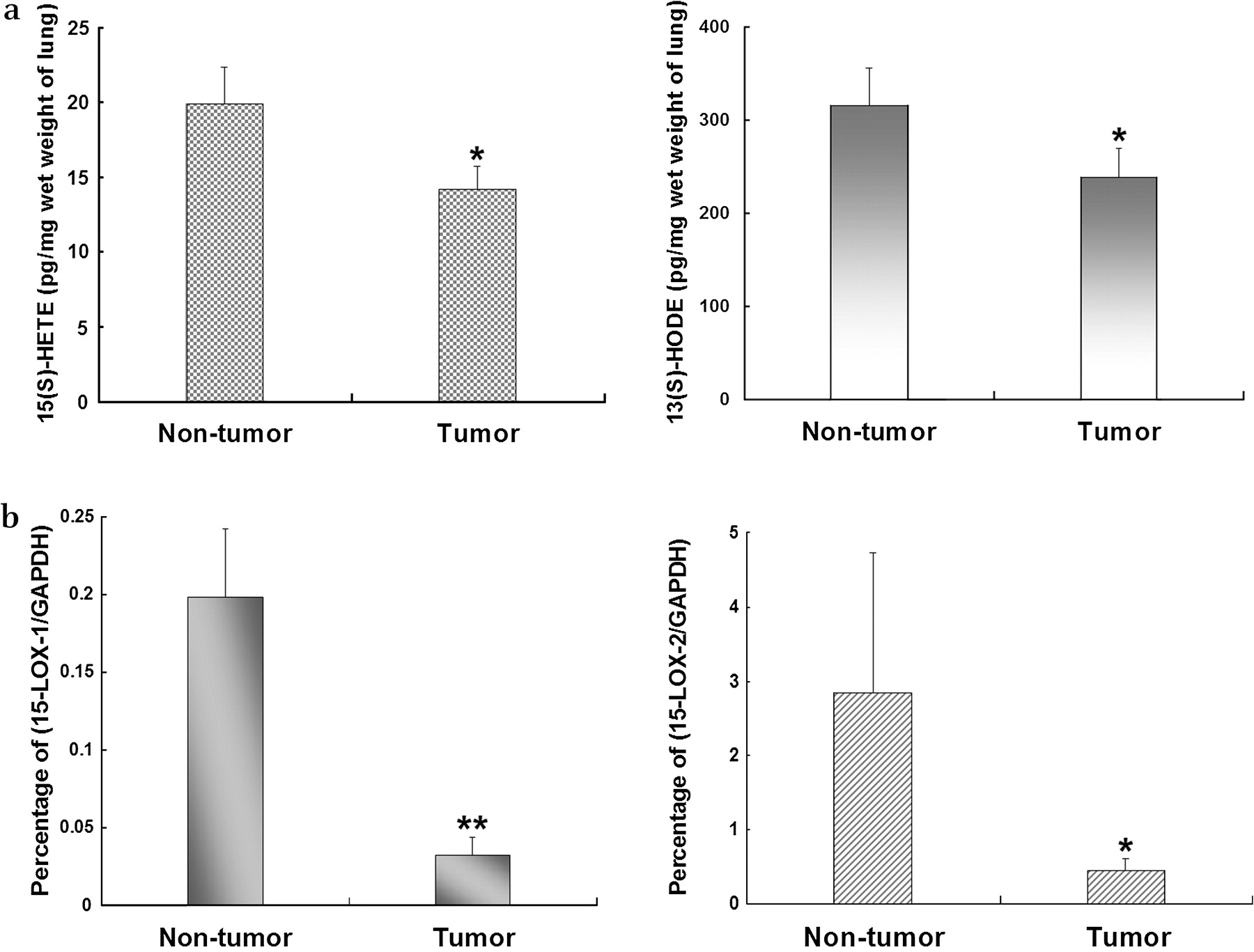
Concentrations of (A) 15-S-hydroxyeicosatetraenoic acid (15(S)-HETE) and 13-S-hydroxyoctadecadienoic acid (13(S)-HODE) and (B) metabolites of 15-lipoxygenase (15-LOX-1 and 15-LOX-2) in human lung tissue. Concentrations of 15(S)-HETE and 13(S)-HODE were measured using ELISA kits and expressed as pg/mg wet weight lung tissue. To determine levels of 15-LOX-1 and -2, total RNA was extracted from lung tumour samples and matching non-tumour lung tissue and the mRNA levels of 15-LOX-1 and 15-LOX-2 were determined by real-time qRT-PCR. All data were normalised by GAPDH and the percentage of 15-LOX to GAPDH was calculated. Data shown as mean±SE. *p<0.05, **p<0.01.
Real-time qRT-PCR was performed to detect mRNA levels of 15-LOX-1 and -2 in human lung non-tumour and tumour tissue. All the data were normalised to the housekeeping gene GAPDH. Compared with non-tumour samples, human lung tumours had significantly lower levels of 15-LOX-1 and 15-LOX-2 (figure 1B).
Development and progression of lung tumours in A/J mice
The time course of lung tumour development was determined in A/J mice following treatment with NNK. Groups of animals were killed starting 26 weeks after carcinogen treatment and every 4 weeks thereafter, with the last time point at 38 weeks after carcinogen exposure. The left lungs of all animals were grossly inspected. At week 38, about 63% of mice treated with NNK had obvious lung tumours, whereas all mice in the control group appeared to be tumour-free. The result of the gross inspection was later pathologically confirmed. Examples of lungs in NNK-treated mice controls are shown in figure 1A and B in the online supplement. Normal lung tissue from non-NNK-treated mice (figure 1C–E in online supplement) and tumour lung tissues separated from non-tumour tissues of the NNK-treated mice (figure 1F–H in online supplement) were stained with H&E to observe the morphological differences between them. Under lower magnification (50×), the pulmonary parenchyma was replaced by tumour tissue which contained abundant blood vessels (figure 1F in online supplement). Under higher magnification (100×, 400×), large deep-stained chromatin granulated nuclei and pathological karyokinesis were observed (figure 1G and H in online supplement). The number of histological lesions in the four sections of the left lung was counted. The number of mice with tumours and the number of tumours in the lungs gradually increased after week 26 (table 3). A significant difference in the number of tumours between NNK-treated mice and controls was not observed until week 34.
Incidence of lung tumours in the left lung following treatment of A/J mice with NNK or PBS
Levels of 15(S)-HETE and 13(S)-HODE during carcinogenesis in lungs of A/J mice treated with NNK
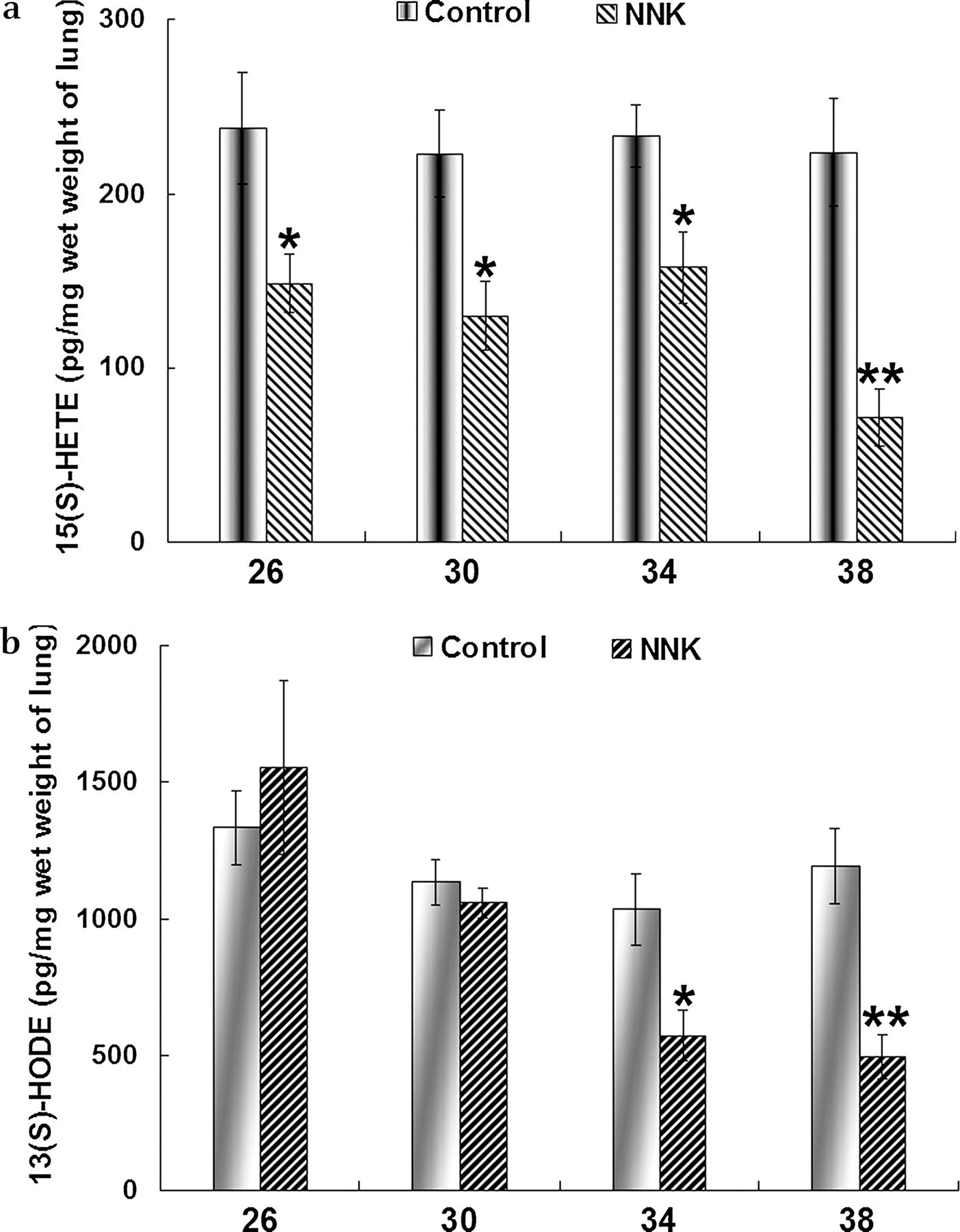
To confirm whether the reduction in 15(S)-HETE and 13(S)-HODE in human lung cancer also occurs in the NNK-mediated lung tumour model, we measured the levels of 15(S)-HETE and 13(S)-HODE in mouse lung tissues by ELISA (figure 2). Compared with the control group, the 15(S)-HETE level was reduced in NNK-treated mice during the period of observation (weeks 26–38 after NNK treatment; figure 2A). The difference in 15(S)-HETE levels between control mice and those treated with NNK was statistically significant at week 26, 4 weeks earlier than the significant appearance of tumour. The 13(S)-HODE level was decreased 30 weeks after NNK treatment and the reduction became significant thereafter (figure 2B).

Concentrations of (A) 15-S-hydroxyeicosatetraenoic acid (15(S)-HETE) and (B) 13-S-hydroxyoctadecadienoic acid (13(S)-HODE) in lung tissue of control and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK)-treated A/J mice at different time points (expressed as pg/mg wet weight lung tissue). Data shown as mean±SE. *p<0.05, **p<0.01.
Expression of 12/15-LOX during lung carcinogenesis in A/J mice treated with NNK
12/15-LOX is the murine orthologue to human 15-LOX-1. The expression of 12/15-LOX was determined at both mRNA and protein levels. The mRNA levels of 12/15-LOX were detected by real-time qRT-PCR and all quantitative analysis data were normalised to the housekeeping gene GAPDH. The results showed that mRNA levels of this enzyme were markedly reduced at week 26 in the NNK-treated group compared with the control group, and that the difference between the two groups increased thereafter (figure 3A). To determine the expression of 12/15-LOX protein in the A/J mice model, western blot analysis and immunohistochemical staining were performed on the mouse lung tissue samples and the results supported the mRNA finding of RT-PCR (see figure 3B and online figure 2). The results of immunohistochemistry further showed that 12/15-LOX was heavily expressed in the lung of the control mice and predominantly localised to the airway epithelium cells. 12/15-LOX was abundant in both the cytoplasm and nucleus (see online figure 2). The reduction in 12/15-LOX immunoreactivity occurred in both the cytoplasm and the nucleus of lung tumour cells, but the reduced intensity in the nucleus was more obvious than in the cytoplasm.

Concentrations of 12/15-lipoxygenase (12/15-LOX) in lung tissue of A/J mice measured by (A) real-time qRT-PCR and (B) western blot analysis. The results were normalised to GAPDH and the percentage of 12/15-LOX to GAPDH was calculated (A). Data shown as mean±SE. In western blot analysis (B), β-tubulin was used as equal loading control. *p<0.05, **p<0.01. NNK, 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone.
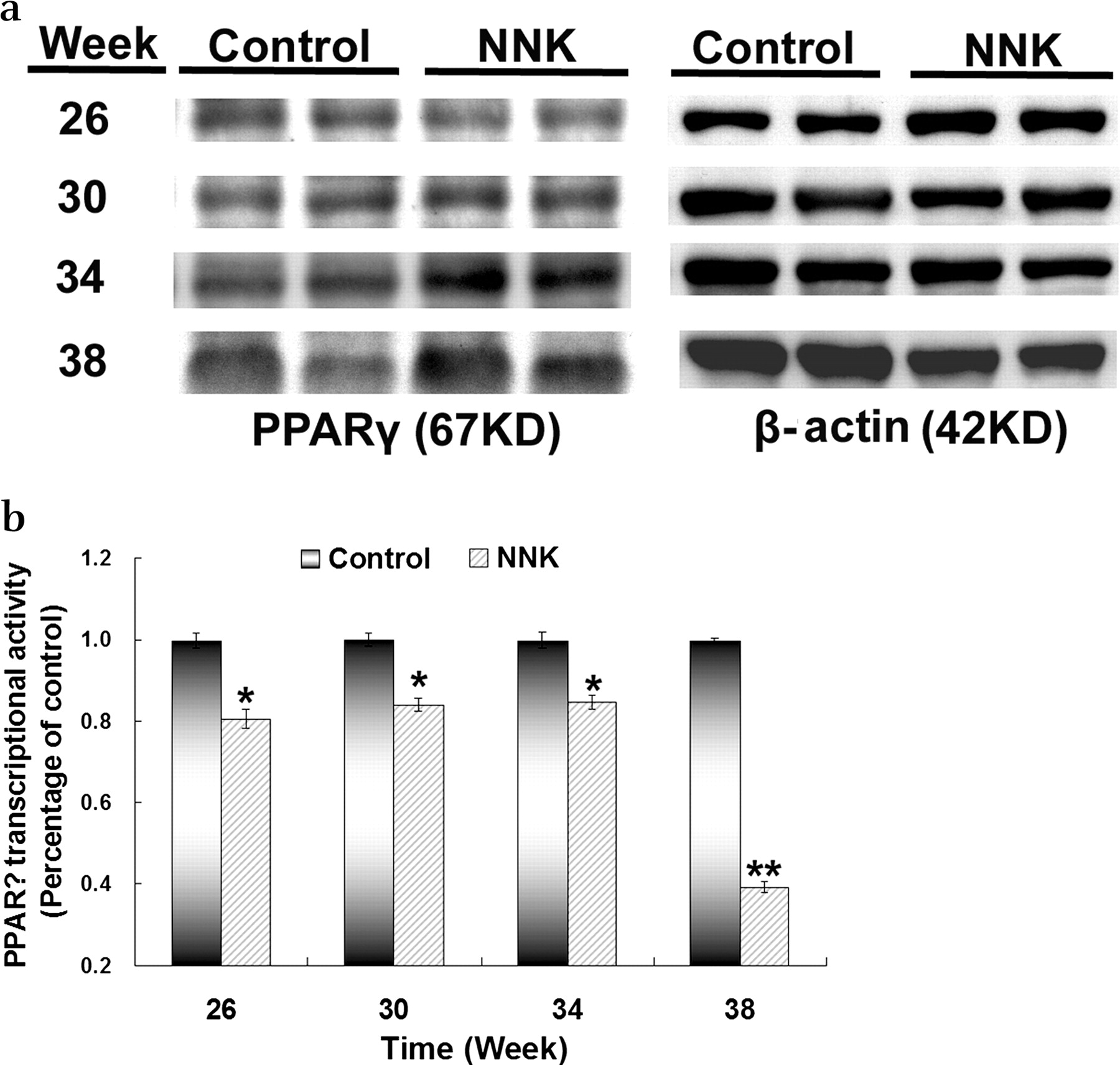
Expression and transcriptional activity of PPARγ during lung carcinogenesis in A/J mice treated with NNK
The expression of PPARγ protein in the A/J mice model was detected by western blot analysis at different time points. The expression of PPARγ increased gradually in the lung tissue in NNK-treated mice from week 30 compared with the control group (figure 4A). Since the expression of PPARγ itself may not reflect its activity, we further analysed PPARγ transcriptional activity. It was found that the activity was significantly reduced rather than increased in lung samples from mice treated with NNK between weeks 26 and 38 compared with lung samples from control mice (figure 4B). Therefore, during the NNK-mediated development of lung tumour in A/J mice, despite the increased expression of PPARγ, its transcriptional activity was reduced and the decreased PPARγ transcriptional activity matched the time frame of the reduction in 12/15-LOX and its metabolite 15(S)-HETE (figures 2 and 3 and online figure 2).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of (A) peroxisome proliferator-activated receptor γ (PPARγ) protein and (B) PPARγ transcriptional activity in 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK)-treated A/J mice and controls. (A) To determine the expression of PPARγ protein, total protein was isolated from the right mouse lung at different time points during lung tumour development and subjected to western blot analysis for PPARγ and β-actin, which was used as a control to confirm equal loading of the amount of protein. (B) To measure PPARγ transcriptional activity, nuclear protein was extracted from mouse lung tissue resected during lung tumour development and measured by PPARγ transcription factor assay (see Methods section). The relative PPARγ transcriptional activity in the lung samples in NNK-treated mice was determined as a percentage of the activity in control mice. Data shown as mean±SD. *p<0.05, **p<0.01.
Discussion
In this study we have shown that the expression of 15-LOX-1 and -2 was significantly decreased in lung tissues of human NSCLC compared with matched non-tumour lung tissue. There is increasing evidence to suggest that 15-LOX-1 and -2 are tumour functional suppressors since their levels are reduced in various types of human cancer,8 11 16 17 and the re-expression of 15-LOX-1 or -2 to restore their enzyme activities has been shown to inhibit tumour growth.10 18–21 Better differentiated NSCLC showed higher 15-LOX-2 expression and there was a significant inverse correlation between 15-LOX-2 expression and tumour grade.22 Furthermore, the methyltransferase inhibitors may induce apoptosis of tumour cells by enhancing the expression of 15-LOX-1,23 indicating that this tumour suppressor gene is inactivated by hypermethylation of its promoter CpG islands. Our finding of a reduction in 15-LOX-1 and -2 in the tumour tissue of NSCLC therefore suggests that 15-LOX-1 and -2 may also function as a tumour suppressor in NSCLC.
In support of the reduction of 15-LOX-1 and -2 in NSCLC, we found that the levels of the 15-LOX metabolites 15(S)-HETE and 13(S)-HODE were significantly reduced in human NSCLC tissue compared with non-cancer tissue. To our knowledge, this is the first evidence of reduced levels of 15(S)-HETE and 13(S)-HODE in human lung cancer, although studies have shown a reduction in 13(S)-HODE and 15(S)-HETE in other tumours.8 11 Studies have shown that both 15(S)-HETE and 13(S)-HODE can function as the endogenous ligand to activate PPARγ.11 24 25 The consequence of the PPARγ activation by its endogenous ligand 15(S)-HETE or 13(S)-HODE is the induction of apoptosis and the arrest of tumour cell growth.8 11 26 The reduction of tumour suppressor genes 15-LOX-1 and 15-LOX-2 may therefore lead to the hypoproduction of 15(S)-HETE and 13(S)-HODE. Subsequently, the low levels of 15(S)-HETE and 13(S)-HODE may fail to activate PPARγ in NSCLC. Surprisingly, in lung cancer the expression of PPARγ is normal or even increased rather than reduced.6 14 This phenomenon has been confirmed in the present study, which showed that the level of PPARγ protein is gradually increased during the development of NNK-induced tumours in the mouse lung. However, normal or even high levels of PPARγ expression do not generate a normal or high activity of PPARγ, which is most likely due to the defect in its endogenous ligands.6 This pathological pattern observed in human lung cancer samples was confirmed in our animal model experiment which showed reduced PPARγ activity in mice with NNK-induced lung tumour compared with controls. This study therefore shows that, during the development of lung tumour, the levels of 15-LOX-1 and -2 together with their metabolites 15(S)-HETE and 13(S)-HODE are reduced and that the activity of PPARγ is decreased, although the expression of PPARγ itself is increased. Such a reverse relationship between 15-LOXs and PPARγ expression has also been found in other epithelial cancers.27
It is unknown when the decrease in the endogenous ligands 15(S)-HETE and 13(S)-HODE occurs and how the change in their levels affects the development of lung cancer. Are the decreased levels of these two ligands early events during the development of lung tumours? Does this change exist throughout the lung tumour development? In order to answer these important questions, we established a lung tumour mouse model to monitor the levels of 15(S)-HETE and 13(S)-HODE during the course of lung tumour development. In this model we used NNK to induce lung tumour in A/J mice. The induced lung tumours were all NSCLC, which is in line with reports showing that lung tumours induced by NNK in mice are almost exclusively adenocarcinomas.28 29 In this model the level of 15(S)-HETE was found to be markedly lower than in control mice after week 26. The level of another major endogenous PPARγ ligand, 13(S)-HODE, was reduced at week 30, 4 weeks later than 15(S)-HETE, and the decrease was significant at weeks 34 and 38 after NNK treatment. In mice, 15(S)-HETE and 13(S)-HODE are mainly metabolised by 12/15-LOX which is orthologous to human 15-LOX-1.30 31 Our animal studies also showed that mRNA and protein levels of 12/15-LOX were reduced in lung tumour tissue of NNK-treated mice compared with normal lung tissue of control mice. The reduction in 12/15-LOX was statistically significant at week 26 and the difference between NNK-treated mice and controls became greater thereafter. This reduction in 12/15-LOX is perfectly in line with the time frame of decreased levels of 15(S)-HETE. It was noted that the difference in the number of lung tumours between NNK-treated mice and controls was not significant until week 34. The marked reduction in 15(S)-HETE and 12/15-LOX therefore predates the significant appearance of lung tumours in our model. These results suggest that the reduction of 15(S)-HETE and 12/15-LOX may have a role in the promotion of lung tumour development. It has been reported that the downregulation of 15-LOX-1 is an early event in the adenoma to carcinoma sequence in the development of colonic and pancreatic cancer.16 17 24 15-LOX-2 can also be an early marker for cancers and is highly expressed in normal prostate glands. In contrast, in PIN (intraepithelial neoplasia) or PIN-like precursor lesions where epithelial cells have lost the single layer organisation and piled up, 15-LOX-2 expression is invariably lost, suggesting that loss of 15-LOX-2 expression may be an early event in the development of prostate cancer.32 15(S)-HETE is one of the major products of 15-LOX-1 and 15-LOX-2.8 9 The reduction in 15(S)-HETE may therefore serve as a potential early marker for human lung cancer, although the carcinogenic role of 15(S)-HETE at the early stage needs to be studied further. It is possible that during tumour development, especially in the early stages, the surrounding normal tissues may provide 15(S)-HETE or 13(S)-HODE via a paracrine mechanism, which may explain the indolence and latency of lung carcinogenesis in some cases.
Similar to human 15-LOX-1, mouse 12/15-LOX is also known to regulate cell growth and death. For example, inactivation of glutathione peroxidase 4 induces apoptosis in a 12/15-LOX-dependent manner in mouse embryonic fibroblasts.33 The reduction in 12/15-LOX is believed to enable lung cells to be more resistant to apoptosis, which may facilitate NNK-induced lung tumour development.
Acknowledgments
The authors thank Johnson Yip, Gary Wong, Rocky Ho and Ernest Chak for their technical assistance in animal experiments, real-time PCR and immunohistochemical staining.
References
Footnotes
Competing interests None.
Ethics approval This study was conducted with the approval of the Joint Chinese University of Hong Kong—New Territories East Cluster Clinical Research Ethics Committee, Hong Kong.
Provenance and peer review Not commissioned; externally peer reviewed.