Article Text

Abstract
Background A proteolytic imbalance has been implicated in the development of “classical” chronic lung disease of prematurity (CLD). However, in “new” CLD this pattern has changed. This study examines the longitudinal relationship between neutrophil proteinases and their inhibitors in ventilated preterm infants and their relationship to microbial colonisation.
Methods Serial bronchoalveolar lavage fluid was obtained from ventilated newborn preterm infants. Neutrophil elastase (NE) activity, cell counts, metalloproteinase (MMP)-9, MMP-9/TIMP-1 complex, SerpinB1 concentration and percentage of SerpinB1 and α1-antitrypsin (AAT) in complex with elastase were measured. The presence of microbial genes was examined using PCR for 16S rRNA genes.
Results Statistically more infants who developed CLD had NE activity in at least one sample (10/20) compared with infants with resolved respiratory distress syndrome (RDS) (2/17). However, NE activity was present in a minority of samples, occurring as episodic peaks. Peak levels of MMP-9, MMP-9/TIMP-1 complex, percentage of AAT and SerpinB1 in complex and cell counts were all statistically greater in infants developing CLD than in infants with resolved RDS. Peak values frequently occurred as episodic spikes and strong temporal relationships were noted between all markers. The peak values for all variables were significantly correlated to each other. The presence of bacterial 16S rRNA genes was associated with the development of CLD and with elevated elastase and MMP-9.
Conclusion NE activity and MMP-9 appear to be important in the development of “new” CLD with both proteinase and inhibitor concentrations increasing episodically, possibly in response to postnatal infection.
- Chronic lung disease of prematurity
- elastase
- alpha-1-antitrypsin
- metalloproteinases
- serpinB1
- proteinases
- bronchopulmonary dysplasia
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/2.0/ and http://creativecommons.org/licenses/by-nc/2.0/legalcode.
Statistics from Altmetric.com
- Chronic lung disease of prematurity
- elastase
- alpha-1-antitrypsin
- metalloproteinases
- serpinB1
- proteinases
- bronchopulmonary dysplasia
Introduction
Chronic lung disease of prematurity (CLD, also called bronchopulmonary dysplasia, BPD) causes ongoing respiratory morbidity and mortality in preterm infants1 2 and is associated with large oversimplified alveoli suggesting aberrant lung development (sometimes called “new” CLD/BPD).3 Inflammation is a hallmark in the development of CLD, with neutrophils and macrophages releasing reactive oxygen species and proteolytic enzymes including the serine proteinase neutrophil elastase (NE)4 and matrix metalloproteinases including MMP-9.5 6 Normally, any free proteinase is effectively neutralised by proteinase inhibitors. Tissue inhibitors of matrix metalloproteinases (TIMPs) are regulators of MMPs, which for MMP-9 in the lung is primarily TIMP-1.5–7 For NE, the best characterised is α1-antitrypsin (AAT) with which it forms an irreversible covalent 1:1 complex; other proteinase inhibitors include secretory leukoproteinase inhibitor (SLPI), elafin, α2-macroglobulin and SerpinB1 (originally called monocyte neutrophil elastase inhibitor) which rapidly binds covalently with NE. SerpinB1 has been studied in rat and mouse models and in newborn baboons but not in human preterm infants.
Early studies reported proteolytic imbalance in preterm infants developing classical CLD.8–10 “New” CLD shows a different pattern with elastase activity being less frequently reported.11–15 An imbalance between MMP-9 and TIMP-1 has also been described, but the importance of MMP-9 in the development of CLD remains unclear.5–7 16–18
The aims of this study were (1) to serially measure NE, AAT, SerpinB1, MMP-9 and MMP-9/TIMP-1 complex in bronchoalveolar lavage (BAL) fluid of preterm infants at risk of developing CLD; (2) to examine the relationships between these variables and their relationship with CLD development; and (3) to correlate enzyme dysregulation with bacterial colonisation as identified by the presence of bacterial 16S ribosomal RNA (rRNA) genes.
Methods
Patient groups
Three groups of mechanically ventilated infants were studied: (1) preterm infants (<32 weeks gestation) who developed mild, moderate or severe CLD using National Institute of Child Health criteria1; (2) preterm infants who developed and recovered from neonatal respiratory distress syndrome (RDS); and (3) a control group comprising term infants who required ventilation for non-respiratory reasons needing ≤28% oxygen throughout the study period. Participants were recruited from the neonatal unit at the University Hospital of Wales, Cardiff between May 2005 and December 2006.
Bronchoalveolar lavage
BAL was performed as previously described.19 20 BAL fluid was collected daily for the first week of life then twice a week until 28 days of age or until extubation. Two aliquots of 1 ml/kg normal saline were instilled and suctioned immediately. Samples were centrifuged at 1000g for 10 min at 4°C within 30 min of collection. The supernatant was aliquoted and stored at −80°C. Pelleted cells were resuspended in phosphate buffered saline, counted with a haemocytometer and cytospins were stained with Diff-Quik for cell differential counts. The unused cell suspension was recentrifuged and the pellet stored at −80°C.
NE activity
NE activity in BAL fluid was measured by the kinetic conversion of Suc-Ala-Ala-Pro-Val-pNA (Bachem, St Helens, UK), as described previously.11 14 The specificity of free NE activity was verified by neutralisation with recombinant AAT (Arriva Pharmaceuticals, Alameda, California, USA).
AAT analysis
The protein concentration in BAL fluid was determined using a BCA Protein Assay Kit (Pierce, Rockford, Illinois, USA). BAL fluid AAT was analysed by western blot analysis with equal protein load as described elsewhere13 using horseradish peroxidase (HRP)-conjugated goat polyclonal anti-human AAT (AB7635; Abcam PLC, Cambridge, UK). Excess purified NE (0.5 μg) was added to BAL fluid for 30 min at room temperature prior to western blot analysis when immunoreactive native AAT was observed coincident with NE activity to test specificity.
SerpinB1 analysis
SerpinB1 was assessed in equal volumes of BAL fluid by western blot analysis using a rabbit polyclonal anti-SerpinB1 primary and an HRP-conjugated goat anti-rabbit secondary antibody.21 A standard curve (3–30 ng) of recombinant SerpinB1 was included on each gel. SerpinB1 was quantified by densitometry using ImageJ software (http://rsb.info.nih.gov/ij/).
Total MMP-9 and MMP-9/TIMP-1 complex
These were measured by ELISA (R&D Systems, Minneapolis, USA) according to the manufacturer's instructions.
Bacterial 16S rRNA analysis
Cell pellet DNA was isolated using the Qiagen RNA/DNA Mini kit (Qiagen, Crawley, UK). PCR was used to amplify the 16s rRNA gene using the primers 27f (5′-AGAGTTTGATC(AC)TGGCTCAG-3′) and 1492r (5′-TACGG(CT)TACCTTGTTACGACTT-3′).22 23 The integrity of extracted DNA was assessed by amplifying human mitochondrial cytochrome oxidase (HMCO) using specific primers. PCR products were separated on 0.7% agarose gels stained with ethidium bromide and visualised in a BioDoc-IT UV Transilluminator (Ultra-Violet Products, Cambridge, UK).
Sequencing of 16s rRNA genes
Amplicons from samples positive for 16s rRNA genes were purified using a Novagen spin-prep PCR clean-up kit (Merck Chemicals, Nottingham, UK) and sequenced using the BigDye Terminator V3.1 cycle sequencing kit (Applied Biosystems, Warrington, UK) and an ABI Prism 3130xl Genetic Analyzer (Applied Biosystems). Sources of bacterial 16S rRNA genes were determined by amplicon sequence comparison using the BlastN website.
Statistical methods
Statistical analysis was performed using SPSS V.15.0. The Mann–Whitney U test was used for non-parametric data and correlations between groups were performed using Spearman correlation coefficients. The presence of NE or 16s rRNA genes between groups was compared using χ2 tests. p Values of <0.05 were considered statistically significant.
Results
Patients
Forty-two infants of <32 weeks' gestational age were recruited: 20 developed CLD (5 mild, 12 moderate, 3 severe CLD); 17 recovered from RDS; 5 infants died before they could be classified and were excluded from the analysis; there were also 6 term controls (table 1). Two hundred and twenty BAL fluid samples were collected: 130 from the CLD group, 45 from the RDS group, 22 from infants who died and 23 from term controls. The median BAL fluid yield was 50% (IQR 38–63%).
Patient characteristics
Cell counts
Median (IQR) peak cell counts in BAL fluid were statistically greater in infants developing CLD (6.75×106 cells/ml; 3.54–17.9×106) than in the RDS group (2.22×106 cells/ml; 1.05–5.50×106, p=0.002) and controls (0.63×106 cells/ml; 0.156–1.26×106, p<0.001). The moderate/severe CLD group (6.30×106 cells/ml; 3.97–11.3×106) had a statistically greater peak cell count than the RDS group (p=0.005). Neutrophils dominated the cell counts but macrophages and some epithelial cells were observed. Peak neutrophil counts were statistically greater in infants developing CLD (3.36×106 cells/ml; 1.79–9.93×106) than in infants with resolved RDS (1.14×106 cells/ml; 0.65–2.88×106, p=0.005) and term controls (0.38×106 cells/ml; 0.048–0.63×106, p=0.001). Infants with moderate/severe CLD (2.56×106 cells/ml; 1.91–7.13×106) had statistically more neutrophils than the RDS group.
Elastase activity
Free NE activity was detected in 26/198 BAL fluid samples (13%). Statistically more infants who developed CLD (10/20) had free NE activity in at least one BAL fluid sample compared with the RDS group (2/17, p=0.013) or controls (0/6, p=0.027). Three of 5 infants with mild CLD had detectable NE in at least one BAL fluid sample. More infants with moderate/severe CLD had free NE detected in BAL fluid than infants in the RDS group (p=0.035). No statistical difference was noted between the RDS and control groups. No sample from the term controls had free NE activity. NE activity was present in 23/129 (18%) lavages from the infants developing CLD and in 3/46 (7%) samples from infants with resolved RDS.
Detection of NE activity was episodic, with the day of first occurrence varying from days 2 to 26 (median 7 days), and NE activity coincided with increased BAL fluid cell counts (figure 1 and online data supplement). In all cases, NE activity was blocked by the addition of exogenous recombinant AAT (data not shown), suggesting an imbalance between proteinases and proteinase inhibitors.

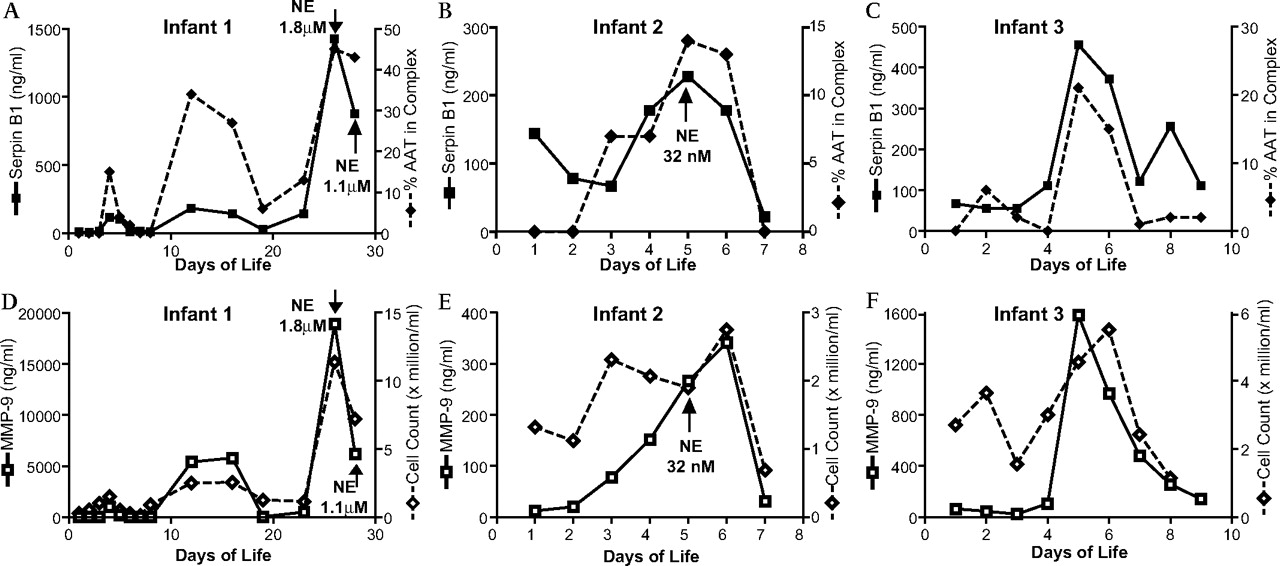
Longitudinal analysis of bronchoalveolar lavage fluid from three infants showing temporal relationships between variables. Graphs A–C show SerpinB1 concentrations (left axis, solid line) and the proportion of α1-antitrypsin (AAT) in complex with serine protease (right axis, broken line) relative to day of life (x-axis). Graphs D–F show the same infants with matrix metalloproteinase (MMP)-9 concentrations (left axis, solid line) and total cell counts (right axis, broken line) against days of life. The arrows indicate the days when free neutrophil elastase (NE) was detected. No free NE was detected in infant 3.
α1-Antitrypsin
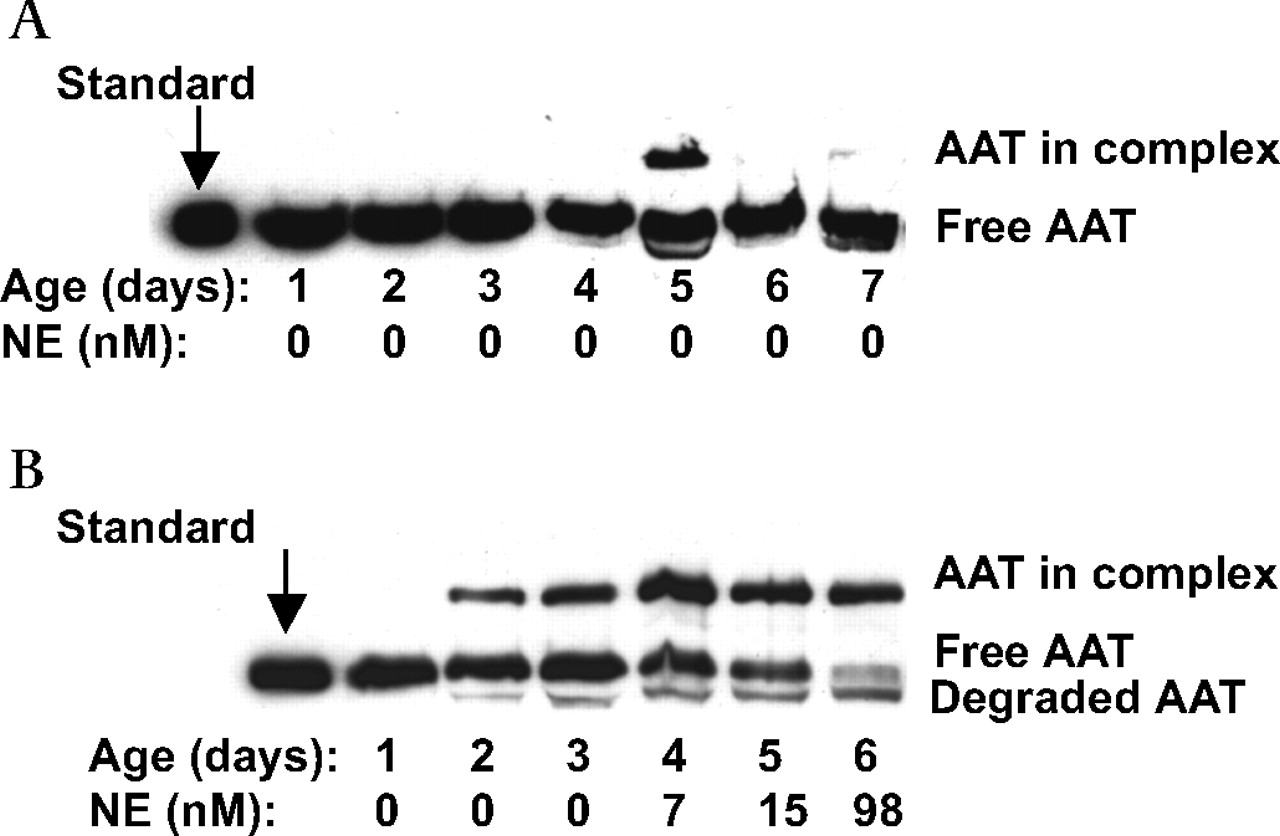
Western blotting detected free AAT as a 53 kDa band, AAT-proteinase complex at 80 kDa and apparent degraded AAT at 48 kDa (figure 2A). AAT-NE complex was detected in 18/20 (90%) infants who developed CLD, which is statistically more than 8/17 (47%) infants whose RDS resolved (p=0.004). The maximum percentage of complexed AAT as estimated by densitometry was statistically greater for infants in the CLD group (median 22%; IQR 13–32%) than for infants whose RDS resolved (median 0; 0–19%, p=0.012) and term controls (median 0; 0–9%, p=0.003), and between the RDS group and infants with moderate/severe CLD (median 22%; 13.5–33.5%, p=0.013).
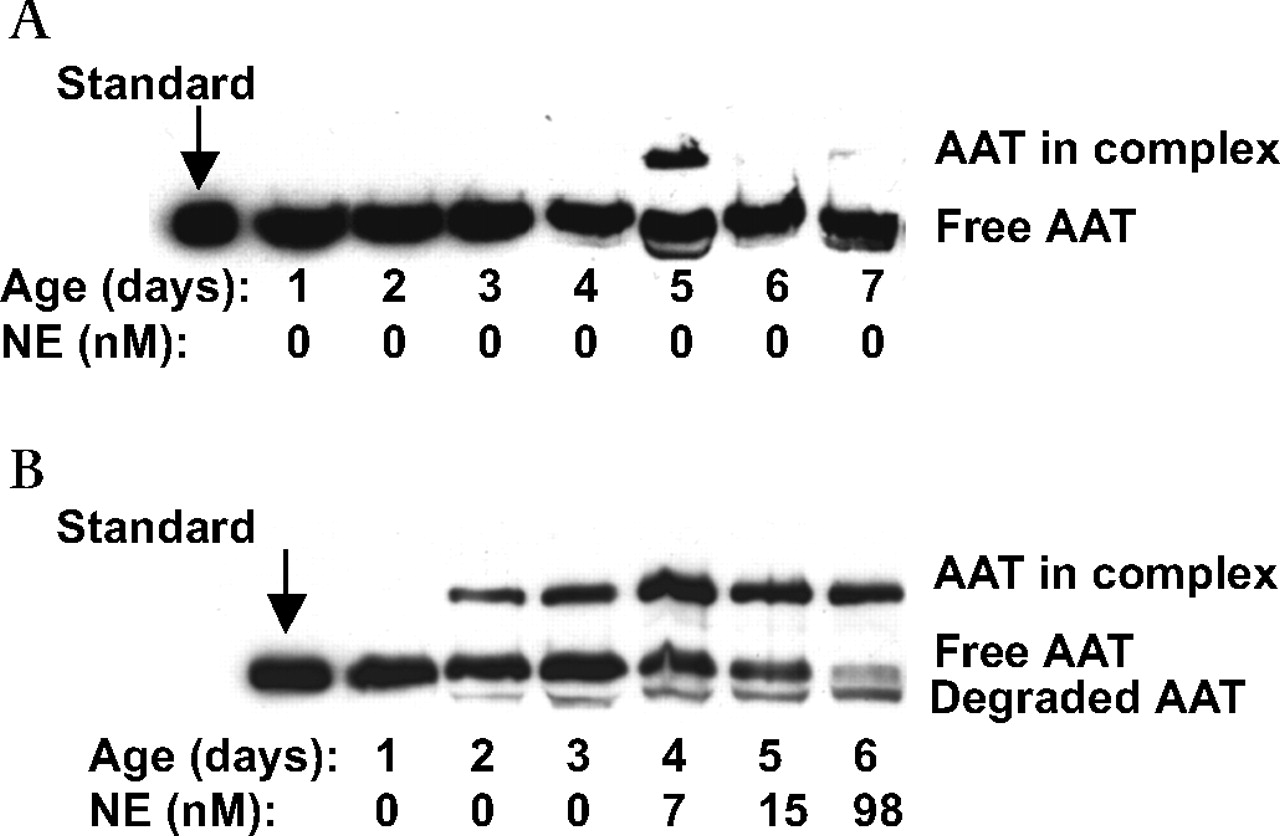
Western blot of α1-antitrypsin (AAT) species in sequential bronchoalveolar lavage (BAL) fluid specimens for (A) a representative infant from the resolved respiratory distress syndrome group in whom active neutrophil elastase (NE) was not detected and (B) an infant from the chronic lung disease of prematurity group in whom active NE was detected. Patient age and NE concentrations are shown below each lane of the western blot. Equal amounts of protein were loaded for analysis and the left lanes show purified plasma AAT migrating at 53 kDa (standard). Additional bands in BAL fluid specimens are the complex with serine proteases at 80 kDa and degraded AAT at 48 kDa. Note that the day 1 samples for both infants contain free AAT only, while day 5 BAL fluid for patient A and days 2–6 for patient B also contain an additional higher AAT-proteinase complex. BAL fluid from days 2–6 of patient B also contains degraded AAT.
Interestingly, BAL fluid collected from some infants with CLD contained both free NE and unbound AAT (53 kDa) (figure 2B). In all cases when a molar excess of NE was incubated with the BAL fluid prior to western blot analysis, the unbound AAT (53 kDa) formed a complex with NE or was degraded.
SerpinB1
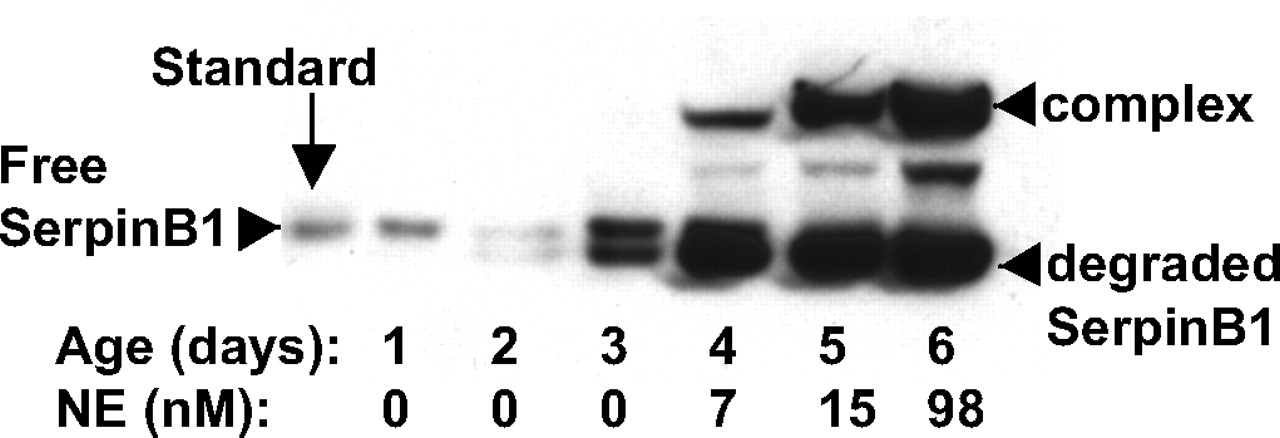
The median (IQR) SerpinB1 concentration on the first day of life was 155 ng/ml (64–414), which did not increase significantly over the first 5 days of life; no statistical difference was noted between the CLD and RDS groups over this period. Median (IQR) peak SerpinB1 concentrations were not statistically different between groups (CLD group: 400 ng/ml; 185–2130; RDS group: 205 ng/ml; 113–480, p=0.161) and term controls (163 ng/ml; 78–164, p=0.107). No statistical difference was seen between peak SerpinB1 concentrations in the RDS group and infants with moderate/severe CLD (455 ng/ml; 191.5–2130, p=0.157). Episodic increases in SerpinB1 were observed more frequently in infants developing CLD than in a minority of RDS infants (figure 3), and these increased SerpinB1 episodes corresponded to episodic increases of cell counts, free NE and NE-AAT complexes (figure 1 and online supplement). Statistically more infants with CLD had covalently complexed SerpinB1 in at least one BAL fluid sample (11/20) compared with infants with resolved RDS (3/17), p=0.020. SerpinB1 complex was not observed in control infants.

Western blot of SerpinB1 species in sequential bronchoalveolar lavage (BAL) fluid specimens of an infant with chronic lung disease of prematurity in whom active neutrophil elastase (NE) was detected. Equal volumes of BAL fluid were loaded for analysis and the left lane shows purified SerpinB1 (10 ng) migrating at 42 kDa (standard). Other species present in BAL fluid are the 66 kDa complex with serine proteinase, a partially degraded intermediate product and degraded SerpinB1 at 38 kDa. A statistically significant increase in both the total concentration and percentage in the complex is seen, occurring with increased NE activity.
MMP-9
Median (IQR) peak MMP-9 protein concentrations were statistically greater in infants who developed CLD (763 ng/ml; 200–1790) than in infants in the RDS group (47 ng/ml; 20–352, p=0.004) and term controls (74 ng/ml; 36–250, p=0.007). Infants with moderate/severe CLD (948 ng/ml; 252–2322) also had statistically greater MMP-9 than those with RDS (p=0.005). Peak MMP-9/TIMP-1 complex concentrations were greater in infants who developed CLD (29.9 ng/ml; 13.1–51.3) than in the RDS group (1.5 ng/ml; 0.9–24.1, p=0.004) and than infants with moderate/severe CLD (29.9 ng/ml; 16.5–51.3) than in infants in the RDS group (p=0.017). Differences between CLD and term infants (7.0 ng/ml; 1.4–24.2) were not statistically significant (p=0.068).
Relationship between variables
Strong correlations were observed between most peak values, particularly MMP-9 and MMP-9/TIMP-1 complexes and NE activity relative to the percentage of complexed SerpinB1 (table 2). In addition, there were strong temporal correlations between cell counts, proteinases and anti-proteinases (figure 1 and online supplement).
Correlation coefficients between maximum values of cell counts, neutrophil elastase (NE) activity, matrix metalloproteinase (MMP)-9 concentrations, SerpinB1 concentrations, percentage α1-antitrypsin (AAT) and SerpinB1 in complex and MMP-9/TIMP-1 concentrations in BAL fluid from ventilated newborn infants
Bacterial 16S rRNA genes
DNA was extracted from 198 BAL fluid samples and satisfactory DNA integrity was confirmed in 162 by HMCO amplification by PCR (only three infants failed to have a satisfactory sample DNA). Bacterial 16S rRNA genes were detected in 75/162 (46%) of these samples; statistically more infants who developed CLD had 16S rRNA genes in their BAL fluid at some point (14/19, 74%) than infants in the RDS group (5/16 (31%), p=0.012) or term controls (2/5 (40%), p=0.155). Statistically more infants with moderate/severe CLD (11/14, 79%) had 16s rRNA genes detected than infants in the RDS group (p=0.001). Elastase activity was found in greater numbers of 16S rRNA gene-positive lavages (19/75, 25%) than 16S rRNA gene-negative lavages (5/87 (6%), p=0.009). Furthermore, elastase activity was higher in the 16S rRNA gene-positive lavages than in the negative lavages (although the median value for both was zero, p=0.0018). The median (IQR) total BAL fluid MMP-9 concentration was greater in 16S rRNA gene-positive lavages (211 ng/ml; 64–1047) than in 16S rRNA gene-negative samples (44 ng/ml; 21–115, p<0.001).
PCR amplicon sequencing showed that the samples with the greatest NE activity and total MMP-9 concentrations contained virulent organisms such as Stegastes aureus and E coli (figure 4A), while the RDS infants with free elastase and the highest MMP-9 concentrations had organisms regarded as being less virulent such as S epidermidis (figure 4B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Scatterplots of (A) peak elastase activity and (B) matrix metalloproteinase (MMP)-9 concentrations for each diagnostic class of infant. Filled symbols indicate 16S rRNA gene-positive specimens and empty boxes indicate negative specimens. Sequencing data for 16S rRNA gene-positive organisms are also shown. Some samples were identified as 16S rRNA gene-positive but no specific microorganism was identified by sequencing. Abbreviations for micro-organisms include S epi (S epidermidis), S haem (S haemolyticus) and P aeru (P aeruginosa). CLD, chronic lung disease of prematurity; RDS, respiratory distress syndrome.
Discussion
This study provides a detailed profile of NE and its inhibitors, AAT (SerpinA1) and SerpinB1 as well as MMP-9 and MMP-9/TIMP-1 complex in ventilated preterm infants. It identified relationships between proteolytic enzymes and their inhibitors and highlights the importance of postnatal infection in proteinase release and in the development of CLD.
Our study is the most detailed longitudinal analysis to date of NE activity in CLD and includes the previously unstudied SerpinB1. Previous studies examined NE at single time points11 13 14 or examined a limited number of samples.12 15 We found that NE activity was episodic and frequently occurred relatively late in the development of CLD, which may cause NE activity to be missed when summary data are presented rather than longitudinally for individual infants. BAL fluid studies of intubated infants prevent comparisons with patients with RDS at equivalent time points. However, our data suggesting that infection may lead to dysregulated inflammation are in agreement with published evidence that prolonged mechanical ventilation commonly results in the development of pneumonia.24
Some infants developed CLD without NE activity being detected. It is possible that NE activity occurred between lavages or following extubation. Also, NE activity may be increased in the pericellular environment as suggested by the presence of complexed AAT and SerpinB1. These complexes, which may be regarded as surrogate markers for regulated NE release, were present in far more samples than was free NE, particularly in infants developing CLD, suggesting successful regulation of proteinases. The simultaneous presence of free NE and unbound AAT (53 kDa) was interesting. This may reflect oxidation of the AAT reactive site methionine residue resulting in a decreased rate of NE-AAT complex formation.25 Thus complex formation occurred after in vitro incubation for a further 30 min with excess NE. Alternatively, increased proteinases including MMP-9 may degrade and inactivate AAT leading to more NE activity. Infection may trigger this dysregulation as it was closely correlated with the increased NE.
SerpinB1 has not been studied in human newborn infants but has been examined in a baboon CLD model.26 Higher NE activity was reported in baboons with “classical” compared with “new” CLD. Baboons developing “new” CLD had 2.5 times the level of SerpinB1 mRNA in BAL fluid compared with term controls, while “classical” CLD was associated with lower SerpinB1 concentrations. We thus hypothesised that a higher SerpinB1 content in “new” CLD may reduce NE activity in human patients and we did find a trend towards higher SerpinB1 levels in infants with CLD than in those with RDS and controls. Temporal relationships were noted between SerpinB1 concentration, active NE and the percentage of SerpinB1 complexed with proteinase, which suggests that the SerpinB1 increase was part of a protective mechanism to regulate NE activity.
AAT is an acute phase reactant.27 Merritt et al8 showed that the AAT concentration almost doubled in lung lavages from infants developing classical CLD. However, we have shown that the SerpinB1 concentration increased locally and rapidly in association with increased NE activity. Moreover, the increase in SerpinB1 was far greater (>10-fold increase for some neonates, figure 3) than has previously been described for AAT8 and may be more important in neutralising NE activity. The mechanisms of SerpinB1 release are unknown as the protein lacks an N-terminal classical secretion sequence.28 The BAL fluid concentration of SerpinB1 correlated well with infiltrating inflammatory cells, particularly neutrophils (which contain the highest concentration of SerpinB1); therefore, SerpinB1 may have been released following cell death or by a non-classical mechanism. However, the possibility of SerpinB1 upregulation and release cannot be discounted.29 Our finding of SerpinB1 increasing in parallel with inflammation is in keeping with those reported in patients with cystic fibrosis where a similar correlation between SerpinB1 levels and neutrophil numbers was also noted.30
The simultaneous episodic increases of neutrophils, proteinases (NE and MMP-9) and anti-proteinases (AAT and SerpinB1) indicate a triggered inflammatory process leading to an increased likelihood of developing CLD; postnatal infection was the most likely candidate to trigger the inflammation. The role of postnatal infection in the development of CLD is recognised31 32 but is frequently underrated. Conventional culture techniques of BAL fluid are confounded by frequent administration of broad-spectrum antibiotics in these patients. Microbial infection or colonisation has previously been confirmed by identification of 16S rRNA genes.33–35 Our finding of 16S rRNA genes detection in patients developing CLD lends further support to postnatal infection being integral in the pathogenesis of CLD. NE was detected more frequently and in higher levels for BAL fluid samples when 16S rRNA genes were detected. This suggests that infection rather than microbial colonisation of the airways is responsible for episodic spikes of protective proteinases and inhibitors.
Previous studies of MMP-9 have reported conflicting results. Dik et al17 showed an increased MMP-9 concentration on days 2 and 4 of life in infants with RDS compared with those with CLD, but Sweet et al5 and Ekekezie et al6 found no statistical difference in MMP-9 levels between groups although both reported a higher ratio of MMP-9 to TIMP-1 in the BAL fluid of infants with CLD. Danan et al7 showed that MMP-9 concentrations, primarily of inactive pro-MMP-9, were increased early in life, but no difference was noted between infants with CLD and those with RDS. Studies of baboons found higher MMP-9 concentrations in extremely premature animals with CLD.16 Our longitudinal study of human infants suggests that MMP-9 may be important in the development of CLD and, as with NE, the increased concentrations were episodic. MMP-9/TIMP-1 concentrations in neonatal BAL fluid have not been reported previously. We found a strong correlation between total MMP-9 and MMP-9/TIMP-1 complex concentrations; however, only a small proportion of total MMP-9 appeared to be in complex with TIMP-1. Furthermore, western blot analysis of our BAL fluid (data not shown) showed that more than half of the MMP-9 was present as dimer rather than monomer, which may interfere with TIMP-1 binding, and the mass of the monomer MMP-9 appeared to be mostly present as the pro-enzyme form. Our finding of an increased MMP-9 concentration in 16S rRNA gene-positive samples suggests that the episodic spikes observed in proteinase concentrations is probably due to infective episodes.
The sequencing data suggest that, while a number of organisms may be associated with high peaks of elastase and MMP-9, those infants who develop CLD had more virulent organisms within their BAL fluid. S aureus and E coli were particularly prominent in infants with CLD with BAL fluid elastase present, while the higher proteinase activity in the BAL fluid of infants whose RDS resolved was associated with S epidermidis, which is frequently dismissed as a commensal.
In conclusion, our study gives a detailed longitudinal profile of NE and MMP-9 and their inhibitors AAT and SerpinB1 as well as MMP-9/TIMP-1 concentrations in the lungs of preterm infants. It showed that episodic peaks of both proteinases and their inhibitors occurred, which may be important in the development of CLD, and we suggest that postnatal infection is a crucial trigger for inflammatory infiltration and proteinase release leading to lung injury resulting in CLD development.
Acknowledgments
We are grateful for the assistance of Jessica Cooley (Immune Disease Institute, Boston, Massachusetts, USA) and colleagues from the neonatal unit in Cardiff, UK. In particular, we would like to thank the parents and infants who participated in this study.
References
Footnotes
Funding Arriva Pharmaceuticals Inc, 1010 Atlantic Avenue, Alameda, CA 94501, USA. Other funders: NIH (HL066548) to ERO and Wellcome Trust.
Competing interests None.
Ethics approval This study was conducted with the approval of the South Wales local research ethics committee and written informed consent was obtained from the parents of study subjects.
Provenance and peer review Not commissioned; externally peer reviewed.