Article Text

Abstract
Background: Granzymes are a subfamily of serine proteases involved in the pathogenesis of many inflammatory disorders. In contrast with granzyme A and B, the role of granzyme K (GrK) in human lung diseases is unknown. Therefore, the release and expression of GrK in allergic asthma, chronic obstructive pulmonary disease (COPD) and bronchopneumonia were investigated.
Methods: Soluble GrK was quantified using an enzyme linked immunosorbent assay in the bronchoalveolar lavage fluid of patients with allergic asthma (before and after segmental allergen challenge), and in patients with mild COPD, pneumonia and in healthy controls. The molecular form of GrK was analysed by western blot. Flow cytometry was performed to determine the cellular expression of GrK.
Results: Compared with healthy controls, there were normal levels of soluble GrK in the bronchoalveolar lavage fluid of patients with COPD, and patients with allergic asthma before allergen challenge. In contrast, soluble GrK was strongly increased in the bronchoalveolar lavage fluid of patients with acute bronchopneumonia. In patients with allergic asthma, there was a significant increase in soluble GrK as well as in GrK expressing CD8+ T cells in the bronchoalveolar lavage fluid 24 h and 72 h after allergen challenge. After allergen challenge, soluble GrK correlated with the percentage of GrK expressing CD8+ T cells. Finally, it was shown that the endobronchial release of the CCR5 ligand CCL3 might be a mechanism for the recruitment of GrK+CD8+ T cells after allergen challenge.
Conclusion: These data provide the first evidence that expression of GrK is upregulated in acute airway inflammation, both in infectious and non-infectious diseases.
Statistics from Altmetric.com
Granzymes (Gr) belong to the family of serine proteases and five of them have already been described in humans, granzymes A, B, H, K and M. They are synthesised by a variety of different cell types (eg, T lymphocytes, natural killer cells, basophils, mast cells) and stored in secretory granules which are located in the effector cell cytoplasm.1 Granzymes have been shown to induce apoptosis of virus infected cells by cleaving intracellular substrates. In addition, they can trigger monocyte activation (GrA) and/or extracellular matrix degeneration (GrA and GrB).2
A role for granzymes A and B in lung diseases such as hypersensitivity pneumonitis (GrA and GrB),3 chronic obstructive pulmonary disease (COPD) (GrA and GrB)4 5 and allergic asthma (GrB)6 7 has been reported. In contrast with granzyme A and B, much less is known about GrK in human diseases. The plasma concentration is elevated during viral infections such as dengue fever and acute cytomegalovirus infection.8 In addition, a functional role for GrK in the development of sepsis was suggested.9
GrK, which has trypsin-like activity,10 is able to induce apoptosis in target cells by cleaving the nucleosome assembly protein SET.11 In addition, it was shown that GrK directly processes the proapoptotic molecule bid (BH3 interacting domain death agonist) leading to mitochondria dependent cell death.12 It was first discovered and purified from lymphokine activated killer cells in which it was less abundant than granzymes A and B.13 In peripheral blood, GrK is mainly expressed by memory CD8+ T cells with a CD27+CD28+CCR5high phenotype and a small population of CD4+ T cells.14 However, there is currently no information about the endobronchial release and expression of this granzyme in human lung diseases. It was the aim of this study, therefore, to investigate the expression and endobronchial release of GrK in patients suffering from allergic asthma, COPD, bronchopneumonia and in healthy controls. In addition, the human asthma model of segmental allergen challenge was used to analyse the regulation of GrK in acute allergic airway inflammation.
METHODS
Subjects
Twenty-two non-smoking patients with mild allergic asthma were included in the study using the following criteria: (1) airway hyperresponsiveness, (2) positive allergen skin prick tests, (3) elevated total or specific immunoglobulin (Ig) E concentrations, (4) dual reaction after allergen inhalation (forced expiratory volume in 1 s (FEV1) fall of >20% of baseline after 5–10 min and >15% after 4–6 h). Inhaled allergen provocation and calculation of the individual provocation dose were performed as described previously.15 Inhaled and segmental allergen challenge were separated by at least 4 weeks. Cromoglycate and corticosteroids were withdrawn at least 7 days before challenge. Inclusion criteria for patients with pneumonia were: (1) onset of dyspnoea and/or cough within the past 7 days prior to bronchoscopy, (2) radiological and clinical signs suggesting pneumonia and (3) enhanced concentrations of C reactive protein in serum (>5 μg/ml). Inclusion criteria for patients with COPD were: (1) smoking history of at least 20 pack-years, (2) dyspnoea on exertion, (3) FEV1/forced vital capacity ratio <70% (GOLD stage I and II) and (4) no use of regular medications. Inclusion criteria for healthy controls were: (1) no smoking history, (2) no history of any respiratory disease and (3) no use of regular medications. All volunteers gave their written informed consent. The study protocol was approved by the local ethics committee.
Segmental allergen challenge
Segmental allergen challenge was performed as described previously.16 Briefly, 2.5 ml of saline were instilled into the left S8 and S4 or S5 segment, and the left S8 was then lavaged using 100 ml of pre-warmed saline. Subsequently, allergen (diluted in 2.5 ml of saline) was instilled into the right S8 and S4 or S5 segment, and the right S8 was lavaged using 100 ml of pre-warmed saline after 10 min. The second bronchoalveolar lavage (BAL) was performed in the left and right S4 or S5 segment, 24 h (n = 15) or 72 h (n = 8) after challenge. For BAL in non-asthmatic volunteers and BAL fluid (BALF) processing, see online data supplement.
Analysis of intracellular and soluble granzyme K
Flow cytometric detection of intracellular antigens was performed as previously described.6 The following antibodies were used: anti-CD3 allophycocyanin (clone MEM-57; ImmunoTools, Friesoythe, Germany), anti-CD4 peridinin chlorophyll protein (PerCP) (clone SK3; BD Biosciences, Heidelberg, Germany), anti-CD8 PerCP (clone SK1; BD Biosciences) and anti-GrK fluorescein isothiocyanate (clone GM24C3; ImmunoTools). The human GrK ELISA was performed as previously described.8
Immunoblotting
BAL samples and recombinant GrK (Alexis Biochemicals, Lörrach, Germany) were mixed with Tris HCl, pH 7.5, containing 1% sodium dodecylsulfate (SDS), 2 mM EDTA and 10% glycerol, and heated at 95°C for 15 min under non-reducing or reducing (by the addition of 5% mercaptoethanol) conditions. Proteins were separated by 10% SDS–polyacrylamide gel electrophoresis and blotted on PVDF membranes (Immobilon-P; Millipore Corp, Bedford, Massachusetts, USA). Blots were incubated with goat anti-granzyme K (C-12; Santa Cruz Biotechnology, Santa Cruz, California, USA) for 2 h at room temperature. For visualisation of antibody binding, membranes were exposed to horseradish peroxidase labelled anti-goat IgG (GE Healthcare, Munich, Germany) and developed using the enhanced chemiluminescence Plus kit (GE Healthcare).
Statistical analysis
Data were analysed using SPSS 14.0. Correlation analysis was performed using Spearman’s correlation coefficient. Comparison of BALF parameters between allergen and saline challenged control segments was performed using the Wilcoxon signed rank test. For the comparison of soluble GrK levels in BALF of patient groups (asthma, COPD or pneumonia) with healthy controls, the Mann–Whitney U test was performed. Probability values of p<0.05 were regarded as significant.
RESULTS
Characteristics of the participants
The characteristics of the patients with asthma are shown in table 1 and allergen induced changes in BALF cell counts are detailed in the online supplement (see table A online). Of the six patients with COPD, four patients had GOLD stage I disease (FEV1 predicted >80%) and two GOLD stage II (FEV1 predicted <80% and >50%). In all patients with bronchopneumonia, there was indirect evidence for a bacterial origin of the infection because the patients responded promptly to antibiotic treatment. Microbiological analysis of BALF in this group yielded positive results in three cases (Mycoplasma pneumoniae, Chlamydia pneumoniae, Legionella pneumophila). Further patient as well as BALF characteristics are detailed in table 2.
Endobronchial concentrations of granzyme K
Soluble GrK could not be detected in BALF from most healthy controls (median 0.0 U/ml, range 0.0–36.6) (fig 1). In BALF obtained 10 min after allergen challenge, no differences in GrK concentrations were detected between allergen challenged (A) and saline challenged control (C) segments (C 10 min: median 4.2 U/ml (range 0.0–13.8); A 10 min: median 5.4 U/ml (range 0.0–17.8)). But there was a significant increase in soluble GrK in BALF 24 h after allergen challenge (C 24 h: median 5.0 U/ml (range 0.0–14.6); A 24 h: median 10.6 U/ml (range 0.0–129.8)), which persisted 72 h after allergen challenge (C 72 h: median 4.8 U/ml (range 0.0–32.0); A 72 h: median 13.9 U/ml (range 3.4–46.2)) (fig 1A). In addition, GrK concentrations in BALF obtained 24 h and 72 h after allergen challenge were significantly elevated compared with healthy controls, while no differences were found between baseline GrK levels in patients with asthma and healthy controls (fig 1A).

Furthermore, in BALF from patients suffering from pneumonia, GrK concentrations were significantly upregulated (median 55.2 U/ml (range 12.6–285.8); p = 0.001). In contrast, patients with mild COPD did not display elevated GrK concentrations in BALF compared with healthy controls (median 1.2 U/ml (range 0.0–15.6); p = 1.000) (fig 1B).
Western blot analysis revealed the presence of free GrK in BALF after allergen challenge and in patients with pneumonia but not in healthy controls. Reducing disulfide bonds with mercaptoethanol substantially increased the amount of free GrK, suggesting that GrK is present in BALF in both forms, the free active and the complexed inactive form (fig 1C).
Correlation of soluble granzyme K with parameters associated with allergic inflammation
Allergen induced elevated endobronchial GrK concentrations were correlated with the severity of the local inflammatory response, as reflected by a significant correlation between soluble GrK and lymphocyte counts (rS = 0.536; p = 0.039), eosinophil counts (rS = 0.619; p = 0.014) and interleukin 5 (rS = 0.581; p = 0.023) in BALF 24 h after allergen challenge. No significant correlation between soluble GrK and soluble granzyme B was found (rS = 0.364; p = 0.183). For concentrations of interleukin 5 and granzyme B and a tabular listing of all p values, see the online supplement (tables C and D online).
Allergen induced increase in BALF GrK+CD8+ T cells
Because GrK is mainly expressed by T cell subsets, we analysed intracellular GrK in CD4+ and CD8+ T cells obtained from BALF 10 min, 24 h and 72 h after allergen challenge (fig 2). Low percentages of GrK expressing CD4+ T cells were detected in all BAL samples which did not change significantly in response to instilled allergen (fig 2B). Ten minutes after allergen challenge, no differences could be detected in GrK expressing CD8+ T cells from allergen challenged and saline challenged control segments (C 10 min: median 28.3% (range 17.8–45.4%); A 10 min: median 25.2% (range 14.8–43.9%)). In contrast, 24 h after allergen challenge, a significant increase in GrK+CD8+ T cells was detected in BAL samples (C 24 h: median 26.1% (range 17.4–40.5%); A 24 h: median 39.7% (range 16.8–62.9%); p = 0.046) which persisted until 72 h after allergen challenge (C 72 h: median 32.7% (range 25.5–51.1%); A 72 h: median 47.8% (range 29.0–76.4%); p = 0.018) (fig 2B). In addition, in BALF, 24 h after allergen challenge, a positive correlation between soluble GrK and GrK expressing CD8+ T cell subsets was observed (fig 2C). No changes in GrK expression in CD4+ and CD8+ T cells from peripheral blood were found 24 h as well as 72 h after allergen challenge (data not shown).

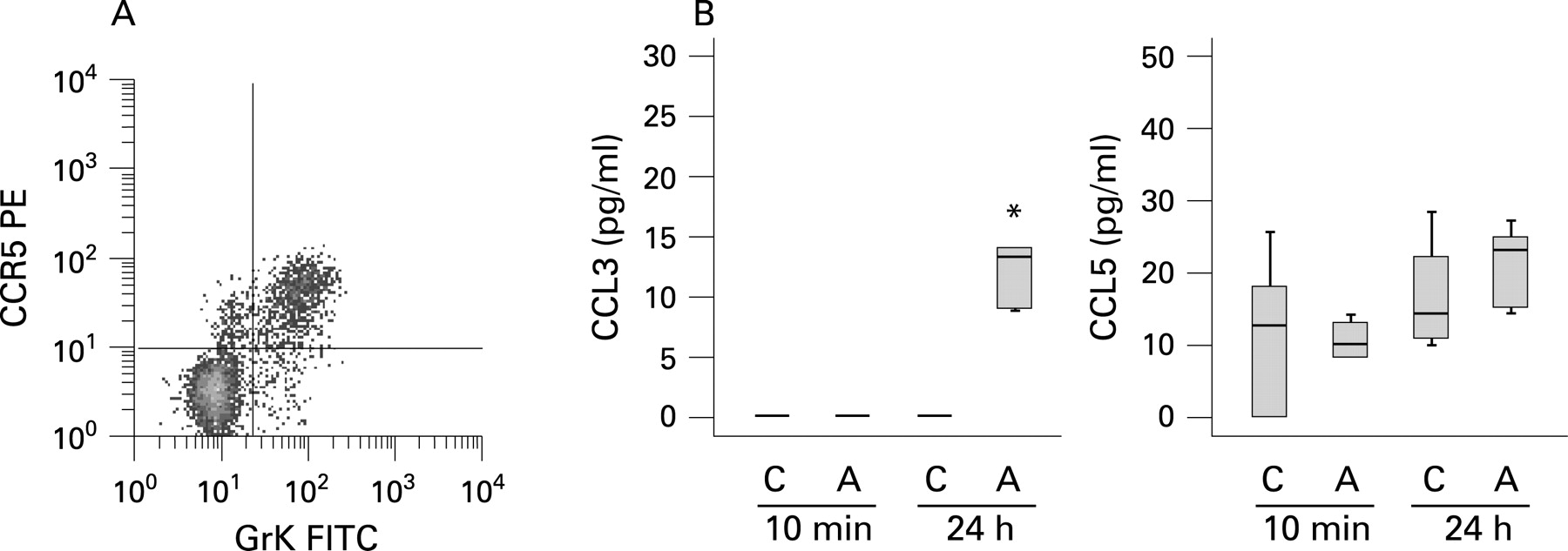
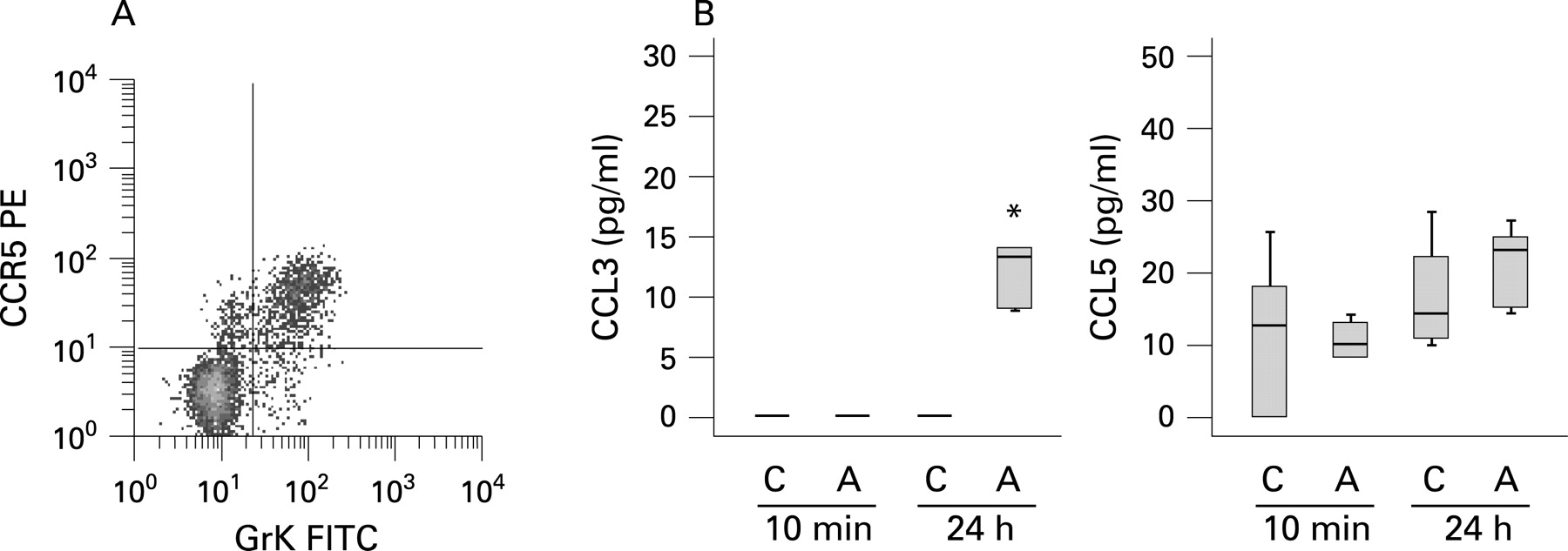
Allergen induced release of CCR5 ligands
Because peripheral blood GrK+CD8+ T cells highly express the chemokine receptor CCR5 (fig 3A)14 and preferentially migrate in response to its ligands CCL3 and CCL5,17 18 we analysed the allergen induced release of CCL3 and CCL5 in BALF. While CCL5 concentrations in BALF were not significantly affected by allergen challenge, a significant increase in CCL3 could be detected 24 h after allergen challenge (fig 3B). In addition, there was a trend for a positive association between allergen specific CCL3 concentrations (A 24 h–C 24 h) and the allergen induced increase in GrK+CD8+ T cells (% GrK+CD8+ T cells from A 24 h - % GrK+CD8+ T cells from C 24 h) which, however, failed to reach statistical significance (rS = 0.627; p = 0.071).
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
In the present study we investigated for the first time expression and release of GrK in human lung diseases. Highest GrK concentrations were found in the BALF of patients suffering from bronchopneumonia. Therefore, this study is the first to report elevated concentrations of GrK in infected airways. Together with other studies which investigated changes in GrK plasma concentrations during viral infection and sepsis,8 9 our data suggest that human GrK is involved in the antibacterial and antiviral host immune response.
In addition, in patients with acute allergic airway inflammation, elevated GrK concentrations were detected which were associated with the severity of the inflammatory response, demonstrating that GrK release is not limited to infections. In contrast, normal GrK levels were found in chronic inflammatory disease patterns as observed in COPD and allergic asthma (without allergen provocation), suggesting that GrK release is a specific event in acute airway inflammation.
Allergen induced GrK concentrations correlate with GrK expressing CD8+ T cells, suggesting that in allergic asthma GrK might originate from these cells. In addition, the low correlation between GrK and GrB concentrations (the latter has previously been reported to be released from basophils in allergic asthma7) suggests that these two granzymes are indeed not secreted by the same cells. At present it is not clear if the allergen induced increase in GrK+CD8+ T cells in BALF is caused by the proliferation of memory CD8+ T cells in the lung or recruitment of these cells from the periphery. The allergen induced proliferation of BAL T cells in vitro has previously been shown.19 In a rat model of asthmatic inflammation, the ovalbumin induced migration of lymphocytes from peripheral blood into the lung has been reported.20
In our study, we demonstrated the allergen induced release of the chemokine CCL3, the ligand of the chemokine receptor CCR5, which is highly expressed on GrK+CD8+ T cells in peripheral blood.14 In mouse studies it has previously been reported that CCL3 is important for CD8+ T cell migration to sites of viral infection and into tumours.21 22 Together, these data suggest that the increase in GrK+CD8+ T cells might be the result of both allergen induced proliferation and recruitment from the periphery through allergen mediated CCL3 release. Notably, we could not find a significant decrease in GrK+CD8+ T cells in peripheral blood 24 h and 72 h after allergen challenge, but recruitment from peripheral blood may be an early event which was not detected at later time points.
The free active form of GrK was present in BALF after allergen challenge, but the functional role of extracellular GrK in the pathogenesis of asthma remains unclear. While for granzymes A and B a variety of extracellular substrates have been reported,2 no such substrates are known for GrK so far. Because a potent inhibitor of GrK, the inter-α-trypsin inhibitor complex, was identified in human plasma,23 it can be speculated that extracellular substrates of human GrK might indeed exist. Regarding activity, GrK is highly related to GrA, suggesting that both granzymes may have similar extracellular substrates and function. For extracellular GrA, a role in lymphocyte migration24–27 as well as in monocyte, fibroblast and epithelial cell activation has already been reported.28 29
In conclusion, we report elevated endobronchial concentrations of human GrK not only in acute bronchopneumonia but also in acute allergic airway inflammation. Allergen induced GrK release was highly associated with GrK expressing CD8+ T cells in BALF. Finally, the allergen induced release of the CCR5 ligand CCL3 suggests that this mechanism might contribute to the recruitment of GrK+CD8+ T cells to the asthmatic lung.
Acknowledgments
The authors thank P Thamm and E Prestin for excellent technical assistance.
REFERENCES
Supplementary materials
web only appendix 63/11/1006
Files in this Data Supplement:
Footnotes
Competing interests: None.
Ethics approval: The study protocol was approved by the local ethics committee.
Linked Articles
- Airwaves