Article Text

Abstract
Background: Whether loss of wakefulness itself can influence pharyngeal dilator muscle activity and responsiveness is currently unknown. A study was therefore undertaken to assess the isolated impact of sleep on upper airway muscle activity after minimising respiratory/mechanical inputs.
Methods: Ten healthy subjects were studied. Genioglossus (GG), tensor palatini (TP) and diaphragm (DIA) electromyography (EMG), ventilation and sleep-wake status were recorded. Non-invasive positive pressure ventilation was applied. Expiratory pressure was adjusted to yield the lowest GGEMG, thereby minimising airway negative pressure (mechanoreceptor) effects. Inspiratory pressure, respiratory rate and inspiratory time were adjusted until the subjects ceased spontaneous ventilation, thereby minimising central respiratory input. Muscle activity during wakefulness, wake-sleep transitions, stable non-rapid eye movement (NREM) sleep and rapid eye movement (REM) sleep were evaluated in the supine position.
Results: In transitions from wakefulness to sleep, significant decrements were observed in both mean GGEMG and TPEMG (1.6 (0.5)% to 1.3 (0.4)% of maximal GGEMG; 4.3 (2.3)% to 3.7 (2.1)% of maximal TPEMG). Compared with sleep onset, the activity of TP during stable NREM sleep and REM sleep was further decreased (3.7 (2.1)% vs 3.0 (2.0)% vs 3.0 (2.0)% of maximal EMG). However, GGEMG was only further reduced during REM sleep (1.3 (0.4)% vs 1.0 (0.3)% vs 1.1 (0.4)% of maximal EMG).
Conclusion: This study suggests that wakefulness per se, independent of respiratory/mechanical stimuli, can influence pharyngeal dilator muscle activity.
- CPAP, continuous positive airway pressure
- CPG, central respiratory pattern generator
- DIAEMG, diaphragm electromyogram
- EMG, electromyography
- EOG, electro-oculography
- EPAP, expiratory positive airway pressure
- GG, genioglossus
- GGEMG, genioglossus electromyogram
- IPAP, inspiratory positive airway pressure
- MTA, moving time average
- NIPPV, non-invasive positive pressure ventilation
- NREM, non-rapid eye movement
- Petco2, end-tidal carbon dioxide tension
- REM, rapid eye movement
- Sao2, arterial oxygen saturation
- Te, expiratory time
- Ti, inspiratory time
- Ti/Ttot, duty cycle
- TP, tensor palatini
- TPEMG, tensor palatini electromyogram
- Ve, minute ventilation
- Vt, tidal volume
Statistics from Altmetric.com
- CPAP, continuous positive airway pressure
- CPG, central respiratory pattern generator
- DIAEMG, diaphragm electromyogram
- EMG, electromyography
- EOG, electro-oculography
- EPAP, expiratory positive airway pressure
- GG, genioglossus
- GGEMG, genioglossus electromyogram
- IPAP, inspiratory positive airway pressure
- MTA, moving time average
- NIPPV, non-invasive positive pressure ventilation
- NREM, non-rapid eye movement
- Petco2, end-tidal carbon dioxide tension
- REM, rapid eye movement
- Sao2, arterial oxygen saturation
- Te, expiratory time
- Ti, inspiratory time
- Ti/Ttot, duty cycle
- TP, tensor palatini
- TPEMG, tensor palatini electromyogram
- Ve, minute ventilation
- Vt, tidal volume
Obstructive sleep apnoea is a common disorder1 characterised by recurrent collapse of the upper airway leading to repetitive episodes of hypoxaemia, hypercapnia and arousal from sleep. This disorder is associated with important consequences for affected individuals including decreased quality of life and likely adverse cardiovascular outcomes.2,3 The pathogenesis of obstructive sleep apnoea is multifactorial, but an anatomically small pharyngeal airway4,5 in combination with decrements in pharyngeal dilator muscle activity during sleep6,7 appear central to the disorder. It is therefore important to understand the effect of sleep on the mechanisms controlling the upper airway musculature.
Pharyngeal dilator muscle activity is influenced by inputs from the chemoreceptors,8 the brainstem central respiratory pattern generator (CPG),9 intrapharyngeal negative pressure10 and possibly vagal input from lung volume.11,12 Studies suggest that most of these mechanisms controlling the upper airway muscles are affected by sleep,13–15 with sleep generally decreasing the sensitivity or responsiveness of the control system. However, it is unclear whether wakefulness alone influences pharyngeal muscle activity or whether its effect on muscle activation is mediated through brainstem respiratory neurons or other chemical/mechanical stimuli.16 At the transition from wakefulness to sleep, there is a decline in genioglossal (GG) and tensor palatini (TP) muscle activity accompanied by a reduction in ventilation and a rise in upper airway resistance.17,18 This decrease in upper airway muscle activity at sleep onset could result from a withdrawal of direct wakefulness stimulation of upper airway muscles, decreased muscle responses to intrapharyngeal negative pressure and/or chemostimuli, or decreased inputs from brainstem respiratory neurons. Most evidence suggests that mechanical (negative pressure) and chemo responses (oxygen and carbon dioxide tensions) are decreased during sleep.13,14 Ventilation falls as well, suggesting decreased CPG output.18 However, whether loss of wakefulness has a direct effect on muscle activity not mediated through other mechanisms has not been tested.
With the application of non-invasive positive pressure ventilation (NIPPV), subjects can be passively ventilated which achieves several goals. First, expiratory positive pressure has been shown to lead to a reduction in the negative pressure reflex thereby minimising mechanoreceptor input to the upper airway muscle.6 Second, by adjusting inspiratory positive pressure and increasing respiratory rate to passively ventilate subjects, NIPPV can eliminate or minimise central respiratory drive to the upper airway muscles as well as to all respiratory muscles such as the diaphragm. NIPPV can also maintain ventilation and lung volume at constant levels. This should allow us to examine upper airway muscle activation during wakefulness, wake-sleep (α–θ) transitions, stable non-rapid eye movement (NREM) sleep and rapid eye movement (REM) sleep without respiratory or mechanical inputs. Thus, pure state (sleep) influences on muscle activity should become apparent.
METHODS
Subjects
Nineteen healthy subjects without any cardiorespiratory or sleep disorder were enrolled into the study. The subjects were aged 19–52 years and had normal body weight (body mass index <27 kg/m2). Six subjects could not be passively ventilated and fall asleep and 3 chose not to finish the protocol, so only 10 subjects completed the entire study. There were no differences between subjects who completed the study and those who did not in terms of age, sex and body mass index. The sample size was based on preliminary data obtained in the first three subjects in the study and a previous study from our laboratory. We determined that 9 subjects would be required, using the change at the α–θ transition with mechanical ventilation to detect a 20% decrease in pharyngeal airway muscle activity with 80% power at the 5% significance level. Written informed consent was obtained from each subject and the protocol was approved by the Human Subjects’ Committee of the Brigham and Women’s Hospital. Women were studied during the follicular phase (days 5–11) of their menstrual cycle as determined by history.19
Equipment and techniques
The subjects wore a nasal mask (Respironics Inc, Murrayville, Pennsylvania, USA) connected to a heated pneumotachometer (Hans Rudolph Inc, Kansas City, Missouri, USA) and a differential pressure transducer (Validyne Corp, Northbridge, California, USA) calibrated with a rotameter for measurement of airflow. Inspiratory (Ti) and expiratory (Te) times were determined from this signal and it was electronically integrated for calculation of tidal volume (Vt). Minute ventilation (VE) was calculated as the sum of all Vt per minute. The duty cycle (Ti/Ttot) was also calculated for each breath. End-tidal carbon dioxide tension (Petco2) was measured from expired air within the nostril using a calibrated infrared CO2 analyser (Capnograph/Oximeter Monitor, BCI, Waukesha, Wisconsin, USA) while arterial oxygen saturation (Sao2) was measured using a pulse oximeter probe attached to the index finger (Capnograph/Oximeter Monitor). Pressure was monitored in the mask with an open catheter attached to a pressure transducer (Validyne Corp). The subjects were instructed to breathe exclusively through the nose. This was ensured by taping the mouth and using an infrared video camera to confirm that the mouth remained closed.
The genioglossal electromyogram (GGEMG) was measured with a pair of unipolar intramuscular electrodes referenced to a single ground, producing a bipolar recording. Two stainless steel Teflon-coated 30-gauge wire electrodes were inserted approximately 12–15 mm into the body of the GG muscle 3 mm lateral to the frenulum on each side using a 25-gauge needle. The needles were removed immediately, leaving the wires in place. The tensor palatini electromyogram (TPEMG) was measured in a similar manner to that of the GG muscle with a pair of unipolar intramuscular electrodes. The tip of the pterygoid hamulus was located at the junction of the hard and soft palate on each side. Using a 25-gauge needle, two stainless steel Teflon-coated 30-gauge wire electrodes were then inserted at a 45° angle along the lateral surface of the medial pterygoid plate to a depth of 10–15 mm into the palate. The needles were then removed, leaving the electrodes in place. This technique has been used previously in our laboratory.8,20 Sucking, blowing and swallowing, which have been shown previously to activate the TP muscle, were performed to confirm placement of the TP electrode.20 The diaphragm electromyogram (DIAEMG) was obtained from electrodes placed at the right 6–8th intercostal spaces adjacent to the costal margin. For all muscles the raw EMG was amplified (Grass Instrument, Quincy, Massachusetts, USA), band-pass filtered (30–1000 Hz), rectified and electronically integrated on a moving time average (MTA) basis with a time constant of 100 ms (CWE Inc, Ardmore, Pennsylvania, USA). Electrocardiographic (ECG) artifacts were removed from the raw DIAEMG using an ECG blanker (CWE Inc). To define maximal EMG for the muscles, subjects were asked to perform several different manoeuvres. They were instructed to (1) swallow; (2) maximally protrude their tongue against the maxillary alveolar ridge; (3) inspire maximally against an occluded tube; (4) suck and blow; and (5) inspire as deeply as possible. Each manoeuvre was performed several times and the maximal EMG recorded during this calibration was designated as 100%. Electrical zero was then determined and each EMG was quantified as a percentage of maximal activation for that subject. The GG and diaphragm muscles are inspiratory phasic muscles so their activation was assessed at two points during basal breathing. The tonic activation was defined as the lowest activation during expiration, the peak activation as the highest activation during inspiration (ie, peak phasic) and the phasic activation as the difference between the peak and tonic activation. However, during mechanical ventilation without spontaneous breathing, no phasic activity was observed during inspiration. Hence, the GGEMG and DIAEMG were reported as the average activation across each breath. As the TP is a tonic muscle without phasic activity, the TPEMG was reported as the average activation across each breath during basal breathing and passive ventilation.
Two EEG channels (C3–A2, O2–A1), left and right electro-oculograms (EOGs) and the submental EMG were recorded to document wakefulness or sleep and score the different sleep stages by a blinded technician. Subjects were maintained in the supine position throughout the study, verified by infrared video camera.
A microprocessor-controlled ventilator (Respironics Inc) that operates in either a continuous positive airway pressure (CPAP) mode or a spontaneous/timed (S/T) mode was used in the study. In S/T mode, this ventilator allowed control of inspiratory positive airway pressure (IPAP), expiratory positive airway pressure (EPAP), respiratory rate, Ti and IPAP rise time. To yield the lowest GGEMG during wakefulness, CPAP mode was applied first and started at 4 cm H2O. This pressure level was increased until the minimal level of GGEMG was obtained or to a maximum of 8 cm H2O. If no obvious reduction in GGEMG was discernible, the subjects were studied on 6 cm H2O. The ventilator was then switched to S/T mode and IPAP was adjusted to obtain the same or a slightly bigger Vt than was observed during eupnoeic breathing. The respiratory rate was set to be the same as or slightly faster than during eupnoeic breathing, and the Ti was set based on respiratory timing in CPAP mode. IPAP rise time was set at 0.05 or 0.1 s. By adjusting the respiratory rate, Ti, IPAP rise time and the pressure difference between IPAP and EPAP, subject passivity—assessed by (a) DIAEMG having no inspiratory phasic activity, (b) GGEMG having no inspiratory phasic activity or pre-activation before onset of airflow, (c) mask pressure showing no reduction before each ventilator-initiated breath, (d) airflow pattern showing a descending ramp shape, and (e) fixed inspiratory-expiratory time ratio—could be achieved (fig 1). Recordings were stopped when there was a departure from this passive pattern until adequate passivity could be achieved or the experiment was terminated.
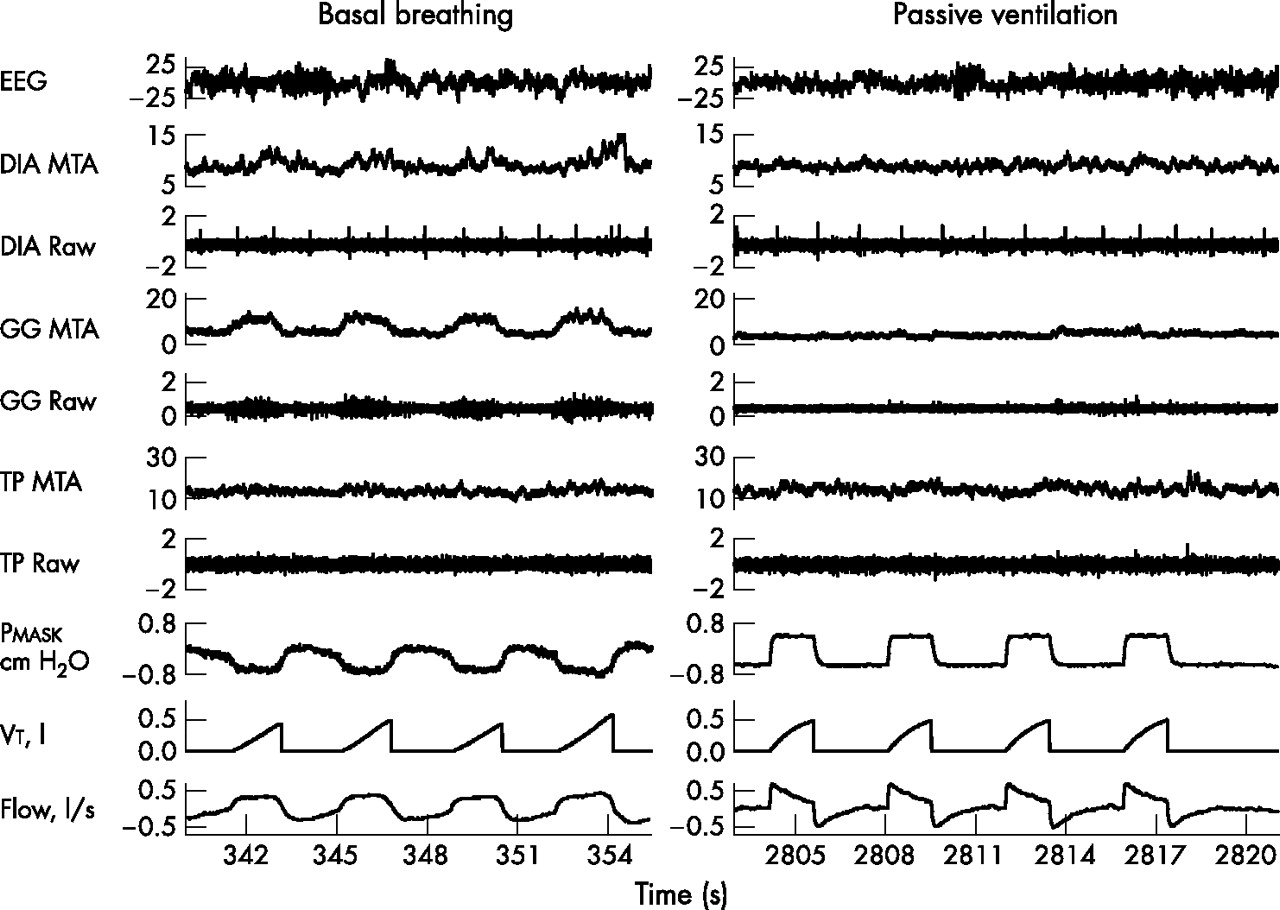
Example of raw data obtained in one individual during basal breathing and passive ventilation during wakefulness. No inspiratory phasic diaphragm or genioglossal activation or pre-activation of the genioglossus occurred before the onset of airflow during passive ventilation. Mask pressure demonstrated no reduction before the ventilator initiated a breath, flow pattern is shown with a descending ramp shape, and fixed inspiratory time/expiratory time ratio all confirm that the subject was passively ventilated without spontaneous active breathing. DIA, diaphragm; EEG, electroencephalography; GG, genioglossus; MTA, moving time average; Pmask, mask pressure; TP, tensor palatini; Vt, tidal volume.
Protocol
Each subject reported to the laboratory at approximately 21:00 h having fasted for at least 4 h. After informed consent was obtained, the sleep staging electrodes and intramuscular EMG wires were placed. Subjects then assumed the supine posture in bed and the nasal mask and pneumotachograph were attached. They subsequently lay with their eyes open in this posture and were allowed to acclimatise to the equipment. Subjects were recorded during both basal breathing and passive ventilation during wakefulness for 5–10 min in each condition. After recording data during wakefulness, the subjects were allowed to fall asleep (in the supine posture) with the mechanical ventilator in place. In order to obtain multiple sleep onsets—primarily transitions from quiet wakefulness to stage 1 sleep—subjects were awoken if they slept for 3–5 consecutive minutes without a spontaneous awakening and then were allowed to fall asleep again. This procedure was repeated until adequate data (at least 3–5 transitions) had been collected. Thereafter, subjects were allowed to fall asleep without interruption. Subjects were subsequently recorded during passive ventilation during stable NREM and REM sleep for 5–10 min in each condition.
Data recording and analysis
All signals (GGEMG, TPEMG, and DIAEMG (raw and MTA), flow, Vt, Petco2, Sao2, EOG, EMG and ECG) were recorded on a computer using an analogue-to-digital converter (1401plus, Cambridge Electronic Design Ltd, Cambridge, UK) and data acquisition software (Spike 2, version 5.03, Cambridge Electronic Design, Ltd). Certain signals (GGEMG, TPEMG, and DIAEMG (MTA), flow, Vt, Petco2, Sao2, EOG, EMG, ECG) were also recorded on a computer using Nihon Kohden software (Polysmith Version 4.0, Neurotronic Inc, Tokyo, Japan). The sample rate varied from 125 Hz for respiratory signals to 1000 Hz for the raw EMG. For each of the three stable conditions (relaxed wakefulness, stable NREM sleep, and REM sleep (phasic and tonic)) during passive ventilation, the mean value for each variable over a 5–10 min recording time was determined.
For each subject, each breath at α–θ transitions was assigned a value of α or θ by visual analysis of the occipital EEG signal by two of the authors (YLL, LD) independently (95% agreement) with any disagreements being resolved. A wake-sleep EEG transition required at least 1 min of α activity followed by clear θ activity lasting for at least 10 s. The transition breaths were defined as α or θ breaths based on having more or less α EEG activity. An adequate α–θ transition was defined as having at least three consecutive α breaths followed by at least two consecutive θ breaths. Each breath in the transition was then assigned a position relative to the transition from −5 to +5, as previously described by Worsnop et al.17 Thus, every transition had breaths −3 to +2, with fewer transitions having breaths in position −5 and −4, and +3, +4 and +5. All ventilation and EMG parameters were calculated on a breath-by-breath basis. If the subject could not be passively ventilated within a defined α–θ transition, the data for the transition were discarded. Thus, all ventilation and EMG parameters were calculated as the mean α level (−5 to −1) and the mean θ level (+1 to +5). Only those subjects having at least 3–5 adequate transitions were used for this analysis.
Analysis of data
All statistical analyses were performed with commercially available software (Excel 2000, Microsoft and SigmaStat + Sigmaplot; SPSS, Chicago, Illinois, USA). A paired t test was used to assess the effect of sleep onset on upper airway dilator muscle activity and ventilation. The Wilcoxon signed rank test was used if data were not normally distributed. Repeated-measures ANOVA with post hoc Student-Newman-Keuls testing was used to assess the effect of stable NREM and REM sleep on pharyngeal airway activity and ventilation. Whenever data were not normally distributed, Friedman’s non-parametric repeated measures comparisons were used. All data are presented as mean (SE) values unless otherwise stated. A p value of <0.05 was considered statistically significant.
RESULTS
Full data sets were acquired in the 10 individuals under all conditions with one exception. In one subject, no adequate TPEMG data were available because of technical problems. Thus, for TPEMG, the data presented are from 9 individuals. For the 10 subjects completing the entire study, the mean (SE) age was 33.0 (4.1) years and the mean (SE) body mass index was 24.0 (0.6) kg/m2.
During basal breathing, inspiratory phasic diaphragm and GG activation were observed in all subjects. Figure 1 shows an example of raw data in one individual during basal breathing and passive ventilation during wakefulness showing no active breathing with NIPPV in place. To achieve passive ventilation, the following mechanical ventilator settings were required: EPAP 6.2 (0.1) cm H2O; IPAP 12.7 (0.4) cm H2O; Ti, 1.2 (0.1) s; respiratory rate 17.7 (0.7) breaths/min. Muscle activity, respiratory data, ventilation, CO2 level and Sao2 during basal breathing and passive ventilation during wakefulness are shown in table 1.
Muscle activity and respiratory data during basal breathing and passive ventilation during wakefulness
Compared with basal breathing, there were significant increases during passive ventilation in Ve and Sao2 and decreases in Ti and Ti/Ttot. No differences in Vt, Petco2 and respiratory rate were observed.
Effects of sleep onset
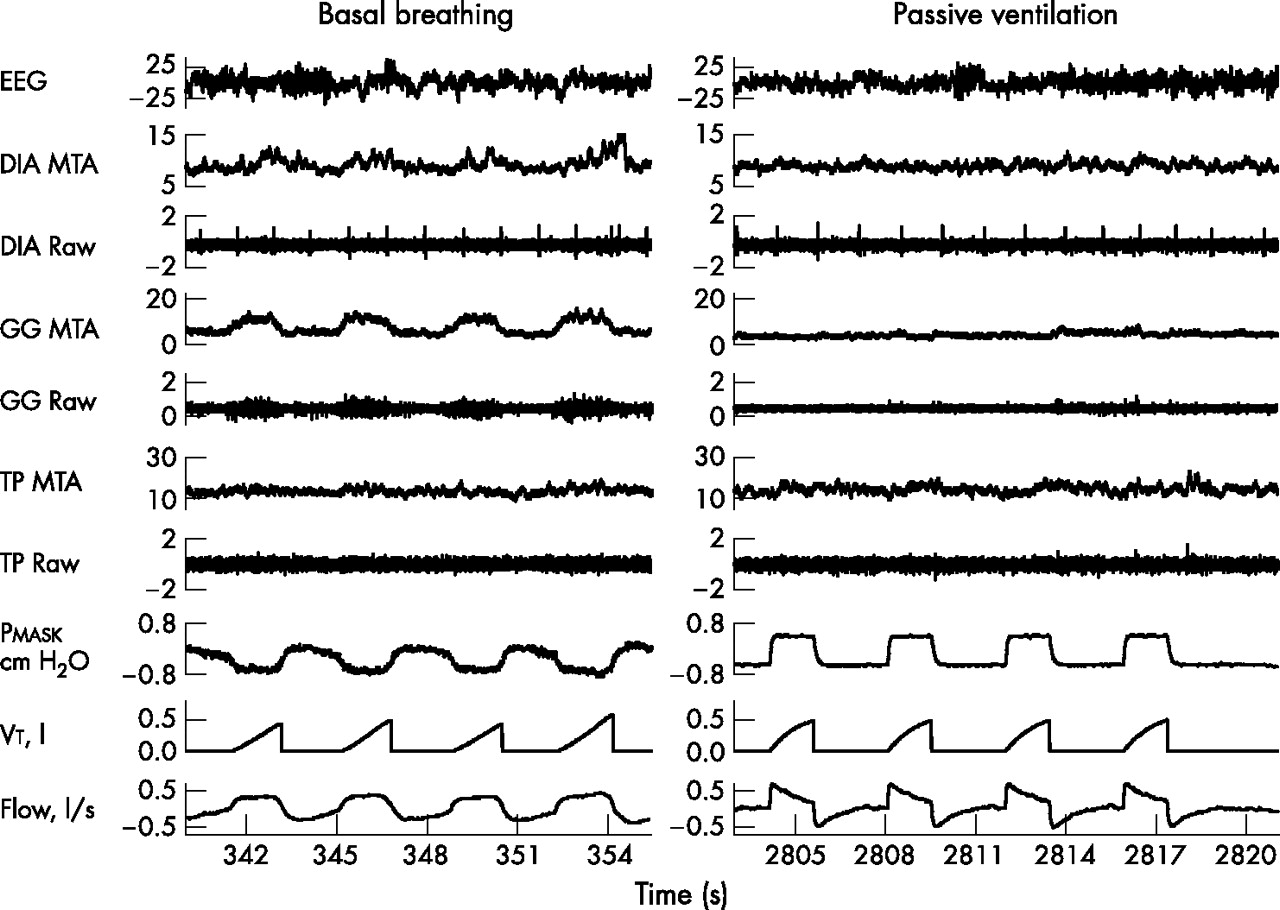
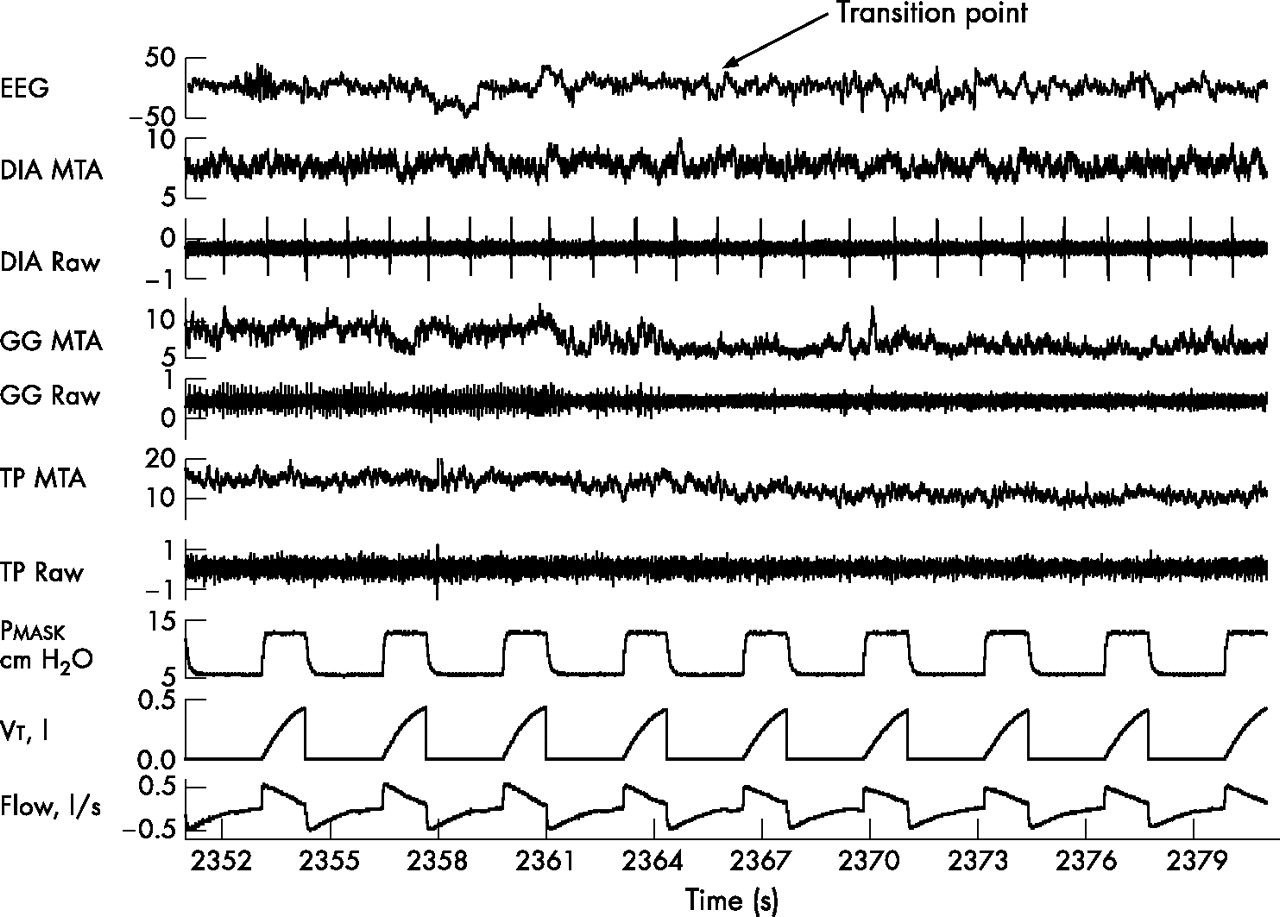
Figure 2 shows an example of a transition from wakefulness to sleep under passive ventilation. For the group, the mean GGEMG and TPEMG during α breaths were significantly higher than during θ breaths (table 2). The individual results and group mean for the GGEMG and TPEMG during α–θ transitions, stable NREM and REM sleep are shown in fig 3A and B.
Muscle activity and ventilation during α–θ transition

Example of a transition from wakefulness to sleep under passive ventilation in one individual. There are substantial decreases in muscle activity in both the genioglossus and tensor palatini across the wake-sleep transition. DIA, diaphragm; EEG, electroencephalography; GG, genioglossus; MTA, moving time average; Pmask, mask pressure; TP, tensor palatini; Vt, tidal volume.
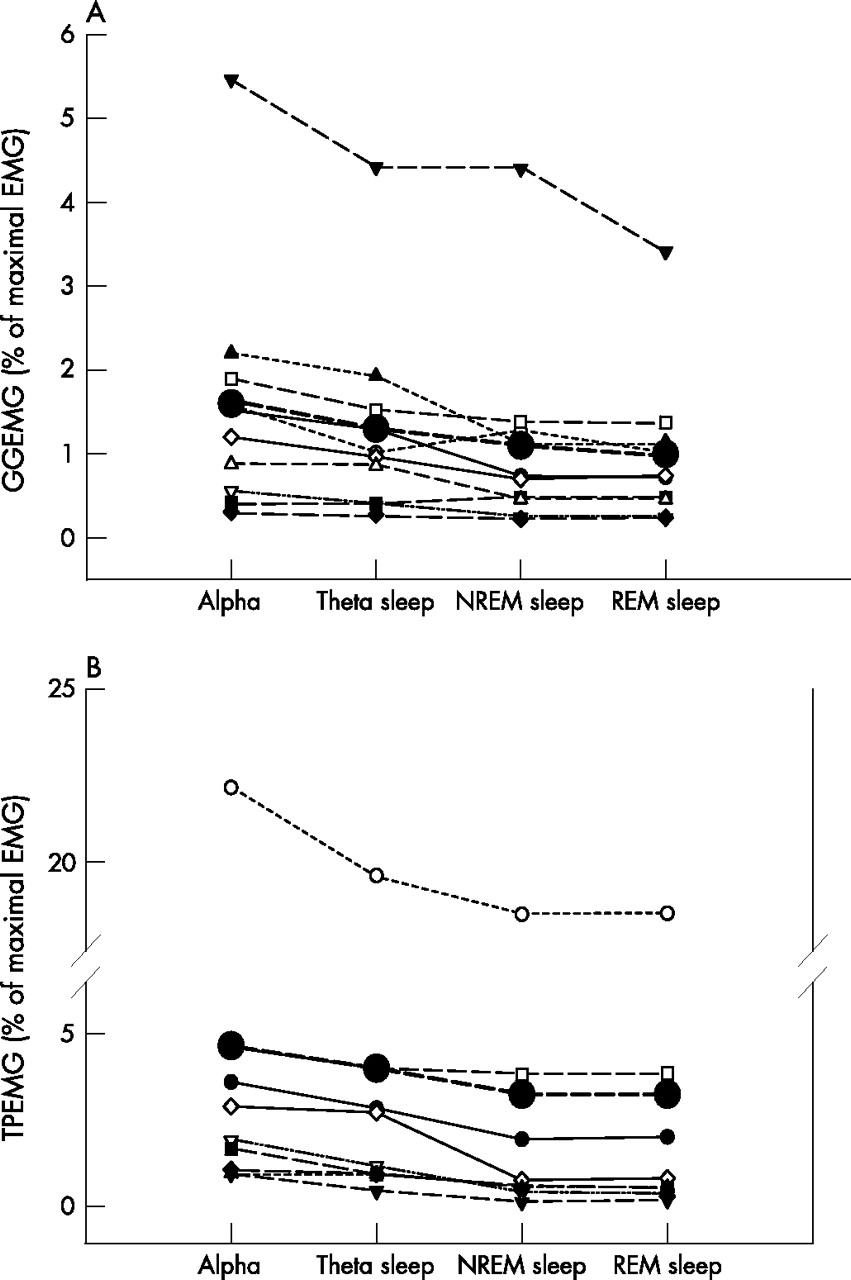
(A) Mean GGEMG and (B) mean TPEMG for each individual during the α–θ transitions, stable NREM and REM sleep. The mean values for each condition are indicated by the large solid circles. GGEMG, genioglossus electromyogram; NREM, non-rapid eye movement; REM, rapid eye movement; TPEMG, tensor palatini electromyogram.
There was no difference in the mean DIAEMG during α versus θ breaths with NIPPV in place. There were also no significant changes in mean Sao2, Petco2, Vt or Ve at α–θ transitions.
Effect of sleep state
Compared with relaxed wakefulness (table 3), there were significant decreases in GGEMG and TPEMG during stable NREM sleep and REM sleep. However, post hoc testing showed that there were no statistically significant differences in GGEMG or TPEMG between stable NREM and REM sleep. DIAEMG did not differ during wakefulness, stable NREM sleep or REM sleep. Other respiratory variables are shown in table 3.
Muscle activity and ventilation during wakefulness, NREM sleep and REM sleep during passive ventilation
Compared with sleep onset (θ breaths), GGEMG was significantly lower during REM sleep but not during stable NREM sleep (1.3 (0.4)% vs 1.1 (0.4)% vs 1.0 (0.3)% of maximal EMG, p = 0.019). However, TPEMG was significantly lower during stable NREM sleep and REM sleep than at sleep onset (3.7 (2.1)% vs 3.0 (2.0)% vs 3.0 (2.0)% of maximal EMG, p<0.001).
DISCUSSION
This is the first study, to our knowledge, to assess the effect of sleep (loss of isolated wakefulness drive) as well as the different sleep states on upper airway dilator muscle activity while minimising the influences of respiratory and mechanical inputs. The results indicate that, at the wake-sleep transition, there are moderate reductions in mean GG and TP muscle activity and, during stable NREM and REM sleep, both GG and TP muscle activity are lower than during relaxed wakefulness. Compared with sleep onset (θ breaths), there were further decreases in the activity of TP during stable NREM and REM sleep but for GG only during REM sleep. Thus, wakefulness has an important independent effect on upper airway dilator muscle activity that is unlikely to be mediated through the respiratory or mechanical control systems.
Immediate effect of sleep on upper airway muscle activity
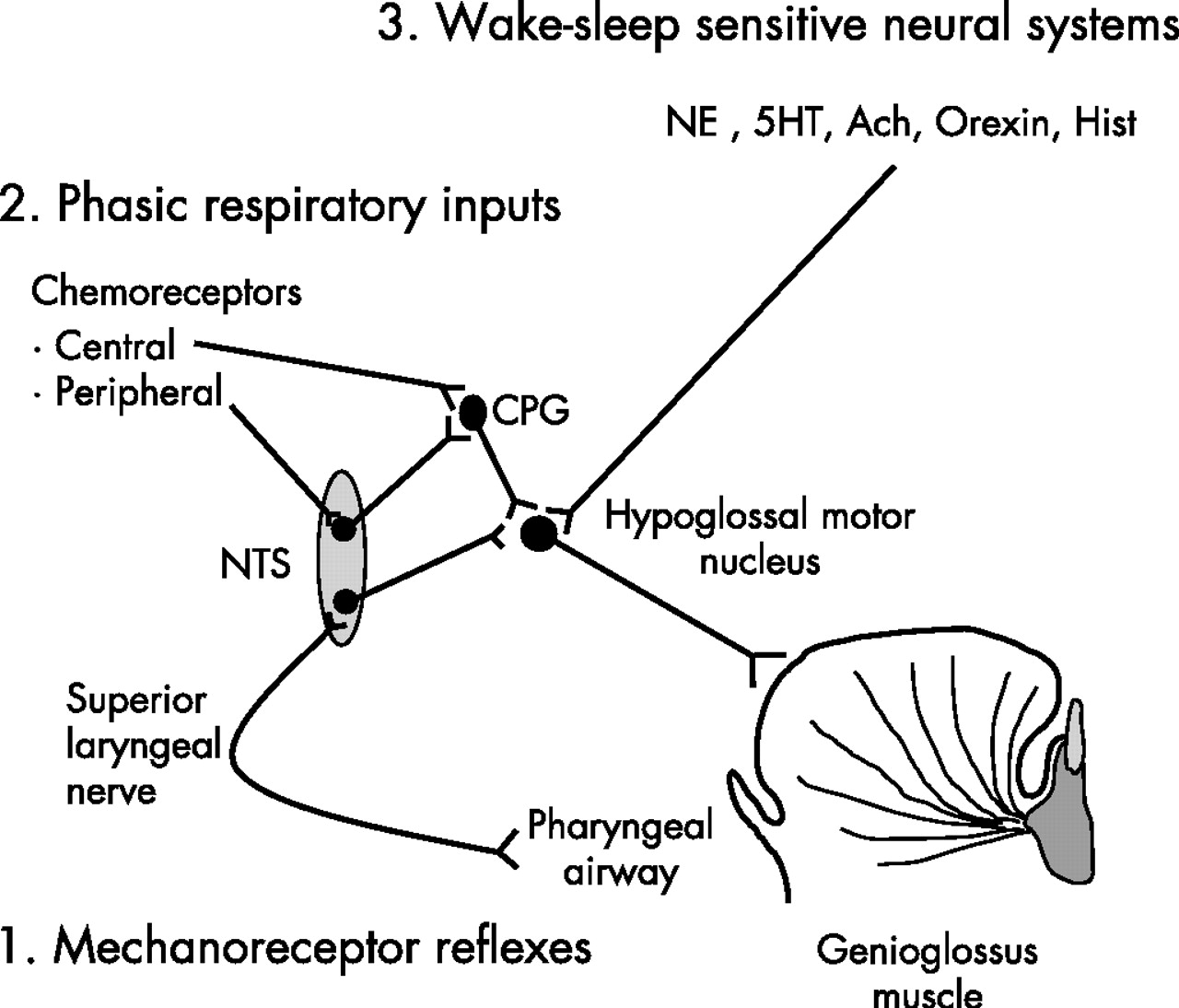
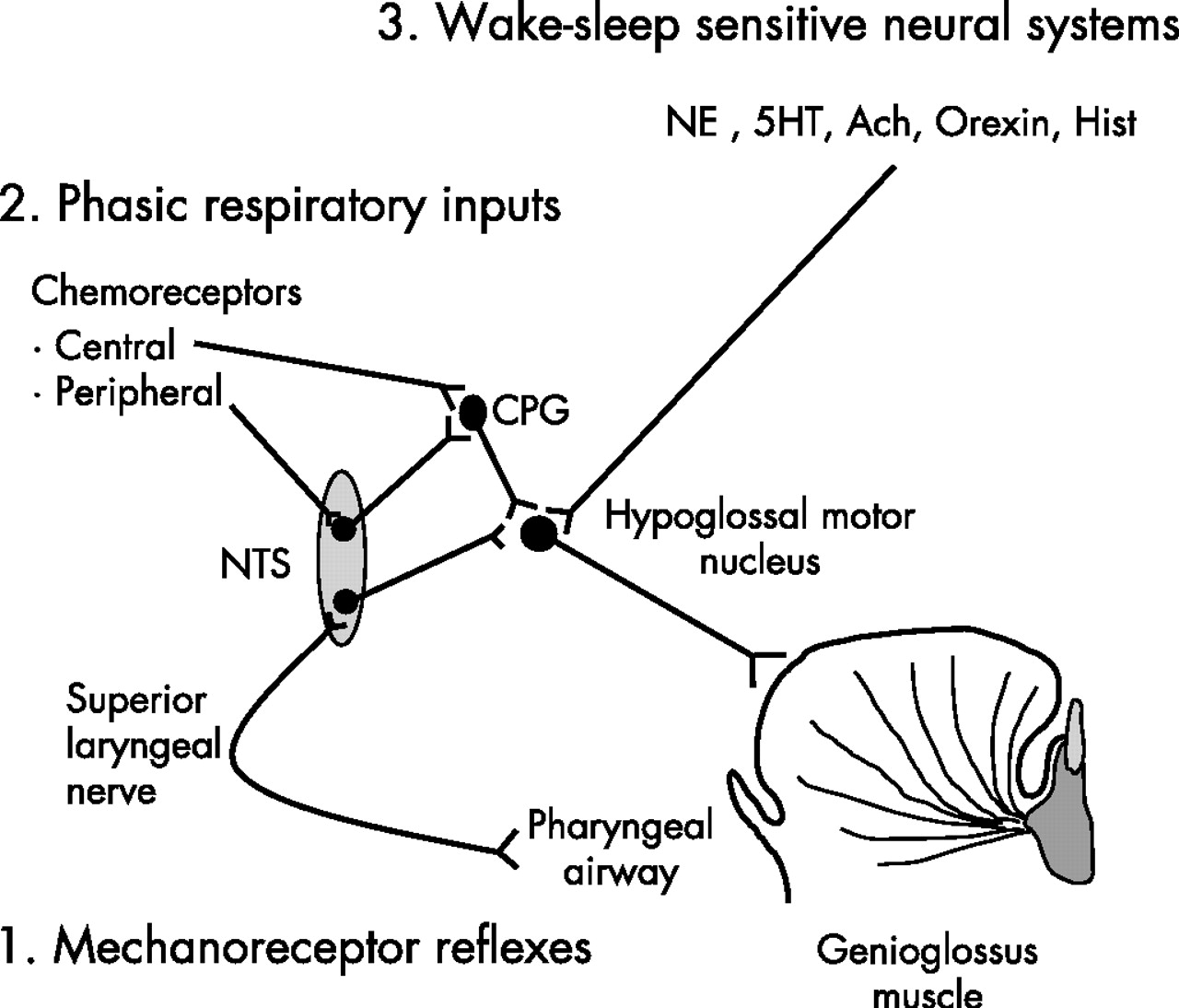
At sleep onset (θ breaths), a consistent decrement in pharyngeal dilator muscle activity has previously been demonstrated by Mezzanotte and Worsnop, together with a reduction in Ve and a rise in upper airway resistance.17,21 However, they observed that there was a subsequent recruitment in muscle activity after the initial fall in GGEMG during the first two θ breaths. By the fifth breath after the transition, GGEMG had largely recovered to stable waking levels. They speculated that GG is recruited in response to increasing negative upper airway pressure and/or rising CO2. By applying CPAP to diminish intrapharyngeal negative pressure, Fogel et al18 reported a similar reduction in GGEMG over the first two θ breaths following a transition, but CPAP prevented the rise in upper airway resistance at the transition and eliminated the subsequent recruitment of GGEMG. It was therefore suggested that the initial reduction in upper airway muscle activity at sleep onset is due to loss of a “wakefulness” stimulus rather than to loss of responsiveness to negative pressure. However, the changes in the upper airway dilator muscle control that occur during sleep onset are complex. Shea et al22 reported that GGEMG responses to brief pulses of negative pressure were minimally reduced (not significantly) in the first five breaths after an α–θ transition compared with stable wakefulness. Nonetheless, it is unclear how chemoreceptive inputs influence GGEMG at sleep onset, as such studies would be difficult to conduct and interpret. Thomson et al15 also reported that, at α–θ transitions, there are prolongations in breath duration and expiratory time in the absence of changes in airway resistance or fluctuations in ventilation and CO2 tension. These observations suggest that there is an abrupt reduction in ventilatory motor output at the wake-sleep transition. As the GG muscle receives inputs from the CPG, it is possible that the change in CPG activity at sleep onset could influence GG muscle activation. Taken together, it is likely that the decrement of GGEMG during sleep onset is a combination of a loss of the wakefulness drive, a mildly decreased negative pressure reflex and a minimally reduced respiratory (CPG) input (fig 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simplified scheme illustrating three potential major sources of neural inputs involved in genioglossus muscle control across wake-sleep states: (1) mechanoreceptor reflexes (negative pressure), (2) phasic respiratory inputs and (3) wake-sleep sensitive neural systems. We believe that (1) and (2) were eliminated by non-invasive positive pressure ventilation. Ach, acetylcholine; CPG, central pattern generator; 5HT, serotonin; Hist, histamine; NE, norepinephrine; NTS, nucleus solitary tract.
In this study, with the application of NIPPV to eliminate or minimise inputs from local mechanical stimuli (negative pressure) and CPG and to maintain ventilation and CO2 levels across wake-sleep transitions, there was a moderate decrement in GGEMG which suggests that the muscle is independently controlled by the wake/sleep state. However, the baseline muscle activity was relatively low which might inflate the percentage change. As a result, the clinical importance of this change in muscle activity is unclear. These observations are also consistent with the concept put forward by Orem and colleagues23 which suggests that there is a wakefulness stimulus to breathing that is not the result of reflexes or sensory inputs. A number of neural systems play an important role in maintaining wakefulness, including noradrenergic, serotonergic, histaminergic, cholinergic, peptidergic and dopaminergic neurons. All demonstrate decrements in the firing frequency from wakefulness to sleep.24 Our studies were designed to determine how sleep-induced changes in several of these neural systems might affect GG activation. Evidence suggests that, during sleep, decrements in the firing frequency of noradrenergic neurons in the locus coeruleus and serotonergic neurons in the raphe could lead to disfacilitation of hypoglossal motor neurons25,26 which innervate GG motor units (fig 4). There is also evidence suggesting that active postsynaptic inhibition of these upper airway motor neurons during sleep via glycine and/or γ-aminobutyric acid may contribute to decrements in muscle activity.27 It is therefore possible that the decrement in GGEMG across wake/sleep transitions observed in this study result from the disfacilitation and/or active postsynaptic inhibition of upper airway motor neurons.
TP muscle activity consistently decreased at the α–θ transition under passive ventilation, confirming previous findings.17,18 Unlike the GG, the TP usually has a primarily tonic pattern of activity without clear phasic respiratory modulation. Neither inspiratory resistive loading during wakefulness and sleep nor the addition of CO2 during sleep20,28 could influence TPEMG. The control of this muscle is therefore substantially different from the GG. However, these studies and ours would suggest that wakefulness has an important influence on the activity of this muscle that is gradually lost across progressively longer sleep times. It is likely that the tonic activity of TPEMG is important for maintaining upper airway patency,7 and the decrement of TPEMG at sleep onset could increase upper airway collapsibility, especially in the area behind the soft palate.
Effect of stable sleep on upper airway muscle activity
Our findings that, with minimal respiratory and mechanical inputs, both GGEMG and TPEMG during stable NREM and REM sleep were lower than during relaxed wakefulness further confirm that the wakefulness drive can modulate upper airway dilator muscle control. However, when compared with sleep onset, both stable NREM and REM sleep led to further TPEMG decrements, but only REM sleep was associated with further GGEMG reductions. This difference could result from the loss of wakefulness input having a different effect on these two muscles. However, it is also possible that our inability to find a difference in GGEMG between sleep onset and stable NREM may have been due to a relative lack of power, given the small number of subjects studied. If such a difference exists, it is likely to be less robust than was seen for TPEMG.
The lack of observed difference in muscle activity between stable NREM and REM sleep deserves comment. Most studies, including protocols in animals and humans, indicate that GGEMG is reduced more during REM than during NREM sleep.29–31 However, one study by Wiegand and colleagues reported no difference in GGEMG between REM and NREM sleep in normal subjects when tonic REM sleep was considered.32 Nonetheless, none of these studies was designed to evaluate the isolated loss of wakefulness on upper airway dilator muscle activity. In this study we observed that, after losing the wakefulness drive to these upper airway dilator muscles, there is consistent tonic muscle activity in both the GG and TP across stable NREM and REM sleep and that this tonic muscle activity is not different between NREM sleep and REM sleep in these two muscles. However, this change in muscle activity may be complicated by a floor effect. Larger changes might have been observed during spontaneous breathing.
Limitations of the study
Several methodological considerations must be examined when assessing our conclusions. First, Sao2 was lower during sleep and there was a trend towards an increased Petco2 level which suggests that all subjects may not have been completely passively ventilated during NIPPV at all times. However, we believe any active ventilation was minimal as our criteria for passive ventilation were strict. We showed that, with NIPPV in place, there must be a loss of phasic GGEMG and a loss of pre-activation (activation before the onset of airflow) in addition to a loss of phasic activation on the surface DIAEMG. The absence of reductions in mask pressure before each ventilator-initiated breath, the fixed inspiratory/expiratory time ratio, and descending ramp shape of the flow pattern also confirmed that there were no spontaneous breathing efforts. We speculate that the difference in lung and chest wall compliance during mechanical ventilation between wake-sleep states might account for this change in ventilation and blood gas levels. This would assume that the lung and chest wall (total respiratory system) becomes less compliant or more stiff during sleep than during wakefulness, which has been carefully demonstrated during anaesthesia33 but it has not been studied during sleep. It is also possible that subjects were simply entrained to the NIPPV rather than passive. However, we believe that the absence of phasic diaphragmatic EMG argues strongly against this. On the other hand, even if subjects were not completely passively ventilated, a very small decrease in respiratory drive cannot explain the large decrement in pharyngeal airway muscle activity from wake to sleep. Second, it is possible that the application of NIPPV may increase resting lung volume and this, in turn, decreases upper airway muscle activity. However, most data suggest that the increased lung volume with increasing expiratory airway pressure is small and stable across wakefulness and sleep.34 Therefore, with NIPPV at fixed pressures across the different states, we believe lung volume would be maintained at a constant level. Third, the application of NIPPV could influence respiration in additional ways including a lowering of upper airway resistance and effects on haemodynamics.35 In addition, the application of positive pressure may directly stimulate positive pressure receptors that have been identified in the trachea, although their function remains unclear. The effects of NIPPV are therefore probably more complex than the simplified view presented here. However, we maintained ventilation and EPAP at fairly constant levels across our sleep states and therefore doubt that these additional effects of NIPPV had a systemic effect on our results or conclusions. Finally, we only studied normal subjects so our observations do not necessarily apply to patients with obstructive sleep apnoea.
In conclusion, our observations suggest that the state of wakefulness has an important influence on pharyngeal dilator muscle activity that is not mediated through respiratory or mechanoreceptive mechanisms. Loss of this wakefulness input may have important consequences for upper airway patency.
REFERENCES
Footnotes
-
Published Online First 27 March 2007
-
This study was supported by NIH grants R01 HL48531, P50 HL60292, NCRR GCRC RR01032 and National Institute of Aging AG024837-01.
-
Competing interests: None.