Article Text

Abstract
Background: Raised concentrations of nitrate and nitrite have been found in exhaled breath condensate (EBC) in airway disease, and it has been postulated that this reflects increased nitric oxide (NO) metabolism. However, the chemical and anatomical origin of nitrate and nitrite in the airways has not yet been sufficiently studied.
Methods: The fraction of exhaled NO at an exhalation flow rate of 50 ml/s (FENO) and nitrite and nitrate in EBC, nasal condensate, and saliva were measured in 17 tracheostomised and 15 non-tracheostomised subjects, all of whom were non-smokers without respiratory disease. Tracheal and oral samples were taken from the tracheostomised subjects and nasal (during velum closure) and oral samples from the non-tracheostomised subjects. Measurements were performed before and after sodium nitrate ingestion (10 mg/kg) and use of antibacterial mouthwash (chlorhexidine 0.2%).
Results: In tracheostomised subjects oral FENO increased by 90% (p<0.01) while tracheal FENO was not affected 60 minutes after nitrate ingestion. Oral EBC nitrite levels were increased 23-fold at 60 minutes (p<0.001) whereas the nitrite levels in tracheal EBC showed only a minor increase (fourfold, p<0.05). Nitrate was increased the same amount in oral and tracheal EBC at 60 minutes (2.5-fold, p<0.05). In non-tracheostomised subjects oral FENO and EBC nitrite increased after nitrate ingestion and after chlorhexidine mouthwash they approached baseline levels again (p<0.001). Nasal NO, nitrate, and nitrite were not affected by nitrate intake or mouthwash. At baseline, mouthwash with deionised water did not affect nitrite in oral EBC or saliva, whereas significant reductions were seen after antibacterial mouthwash (p<0.05 and p<0.001, respectively).
Conclusions: Besides the salivary glands, plasma nitrate is taken up by the lower airways but not the nasal airways. Nitrate levels in EBC are thus influenced by dietary intake. Nitrate is reduced to nitrite by bacterial activity which takes place primarily in the oropharyngeal tract of healthy subjects. Only oropharyngeal nitrite seems to contribute to exhaled NO in non-inflamed airways, and there is also a substantial contribution of nitrite from the oropharyngeal tract during standard collection of EBC.
- EBC, exhaled breath condensate
- FENO, fraction of expired nitric oxide
- CF, cystic fibrosis
- PCD, primary ciliary dyskinesia
- NO, nitric oxide
- dH2O, deionised water
- exhaled nitric oxide
- exhaled breath condensate
- nitrite
- nitrate
Statistics from Altmetric.com
- EBC, exhaled breath condensate
- FENO, fraction of expired nitric oxide
- CF, cystic fibrosis
- PCD, primary ciliary dyskinesia
- NO, nitric oxide
- dH2O, deionised water
Nitric oxide (NO) is present in exhaled air of animals and humans,1 and a number of respiratory diseases characterised by airway inflammation have been associated with altered levels of exhaled NO.2 It is, for example, well known that the exhaled NO concentration is increased in asthma.3 However, the opposite has been shown in studies of exhaled NO in patients with cystic fibrosis (CF)4,5 and primary ciliary dyskinesia (PCD).6,7 The increase in exhaled NO in asthma has been suggested to be due to an increased expression of inducible NO synthase (iNOS) in the bronchial epithelium,8 whereas patients with CF seem to express less iNOS in the airways than healthy subjects.9
In the oral cavity, NO is also formed via bacterial reduction of salivary nitrate to nitrite, mainly in the deep crypts in the posterior part of the tongue and subsequently via further chemical reduction of nitrite to NO.10–12 Thus, nitrite in the upper airways may serve an important host defence purpose, being a substrate for NO formation.13 Also, the nitrite-derived NO in the oropharyngeal tract has been shown to influence NO measurements in exhaled air.11 Although most studies on non-invasive markers of airway inflammation have focused on exhaled NO, there has recently been an increasing interest in measuring compounds found in exhaled breath condensate (EBC).14 Exhaled breath is saturated with water vapour that can be condensed by cooling. Exhaled breath also contains water droplets and available evidence suggests that non-volatile substances can be transported from the respiratory tract dissolved in these droplets.15 Several non-volatile compounds have been reported to be found in EBC, including nitrite and nitrate. These nitrogen oxides, when found in EBC, have been proposed to be oxidised metabolites of NO primarily and, as such, markers for airway inflammation.16 However, the origin of markers in EBC has not yet been sufficiently studied. Furthermore, the increased17 or unchanged18 levels of nitrite in EBC of patients with CF and PCD, respectively, speak against a direct relation between the level of NO in exhaled air and nitrite in EBC.
To elucidate the origin of nitrate and nitrite in EBC and to study the possible formation of nitrite from nitrate and NO from nitrite in the respiratory epithelium of the lower and upper airways isolated from the oropharyngeal tract, NO was measured and EBC collected during tracheal and oral exhalations in patients with a permanent tracheostomy and NO was measured and condensate collected from the nasal airways in non-tracheostomised subjects. These measurements were repeated before and after ingestion of nitrate and mouthwash with an antibacterial agent to manipulate the concentration of nitrite and nitrate in the airways.
METHODS
NO measurements
Exhaled NO measurements (fraction of expired NO at an exhalation flow rate of 50 ml/s; FENO) were made by chemiluminescence according to ATS guidelines.19 The subjects were asked to inhale NO-free air. After inhalation they immediately exhaled with a target flow rate of 50 ml/s against a resistance, giving rise to an oral or tracheal pressure of 8–15 cm H2O, for a period of 6–10 seconds. The exhaled air was led into the Aerocrine NO System or Niox (Aerocrine AB, Solna, Sweden) for measurement of NO. Nasal NO was measured by aspirating air (5 ml/s) from one nostril during a breath hold period of at least 20 seconds and the NO plateau was registered (Niox). Ambient NO levels were below 10 ppb during nasal NO measurements.
Condensate collection
EBC was collected orally or from the tracheostomy (tracheal EBC) using a specifically designed condenser (EcoScreen; Jaeger, Würzburg, Germany). The subjects were asked to breathe at normal frequency and tidal volume while wearing a nose clip. To standardise the collection we used a spirometer with the total exhaled volume set to 20 litres (SpiroPro+; Jaeger, Würzburg, Germany) which took 2–3 minutes. The condenser tube was centrifuged for 1–2 minutes at 400g at +4°C before aliquoting the condensate. To avoid nitrite/nitrate contamination and to improve the Teflon coating on the condenser tubes, we deposited a Teflon-like coating by means of plasma polymerisation on the existing coating (in-house method at Institute for Surface Chemistry, Stockholm, Sweden). The plasma treated condenser tubes were thoroughly cleaned between all condensate samplings in a slightly acidic desinfectant (Descogen; Jaeger) using ultrasound. The absence of contaminating nitrite/nitrate on condenser tube surfaces after this cleaning procedure was confirmed by rotating tubes end over end with 1 ml nitrate-free and nitrite-free deionised water (dH2O) for 15 minutes and then analysing the content of nitrate and nitrite. This resulted in nitrate concentrations of <1 µM and nitrite concentrations of <0.2 µM after cleaning of plasma treated condenser tubes (used in this study), which was not always the case in non-plasma treated condenser tubes after cleaning.
Nasal condensate collection
Nasal condensate collection was introduced as a new technique. The subjects were asked to hold their breath and to voluntarily close the velum for 30 seconds while an air stream with a flow rate of 80 ml/s was led through the nose from one nostril to the other via the introduction of nasal olives and then into the condenser (EcoScreen). This was repeated six times in alternating directions. All condensate samples were immediately frozen and stored at −80°C (⩽3 weeks) for later analysis.
Saliva collection
The subjects were asked to chew on a small piece of plastic for 1 minute and saliva (1–2 ml) was collected in a tube, centrifuged for 10 minutes at 400g at +4°C and the supernatant was stored at −80°C for later analysis.
Analyses
For analysis of nitrite/nitrate and amylase, commercially available kits were used: Nitrate/Nitrite Fluorometric Assay kit; Cayman Chemical Company, Ann Arbor, MI, USA (detection limit 0.2 µM) and EnzChek Amylase Assay Kit; Molecular Probes Inc, Eugene, OR, USA (detection limit 0.1 IU/l corresponding to 0.3 ng/l protein).
Subjects
A total of 17 subjects with a permanent tracheostomy attending the Respiratory Unit at Danderyd Hospital, a national center for tracheostomised patients in Sweden, were recruited for the first two studies (table 1). All patients had tracheal cannula systems allowing them to breathe either via the mouth with the tracheal cannula plugged or via the tracheostomy with the upper airway sealed off. Their respiratory failure was extrapulmonary and none of them had a pulmonary disease. However, in some of the patients the upper part of the trachea was colonised by bacteria such as Pseudomonas aeruginosa, Streptococcus pneumoniae, Staphylococcus aureus, Proteus mirabilis, Streptococcus (B, G) and/or Enterobacter cloacace. Fifteen healthy non-tracheostomised controls aged 24–44 years (seven men) were recruited for the third and fourth study. All subjects were non-smokers and had no history of allergy or chronic pulmonary disease. They did not have any ongoing respiratory infection.
Characteristics of participating tracheostomised subjects
The studies were approved by the local ethics committee and all subjects gave informed consent.
Orally and tracheally exhaled NO in tracheostomised patients
Ten tracheostomised patients had fasted overnight and the experiments were performed in the morning. Baseline values of oral and tracheal FENO were measured using the Aerocrine NO system 30 minutes and immediately before sodium nitrate (10 mg/kg body weight dissolved in 150 ml dH2O) was ingested. This nitrate intake corresponds to approximately 200 g greenhouse lettuce in a 70 kg person.20 The mouth was rinsed with tap water immediately after the nitrate intake. Measurements of FENO were then repeated at 30, 60 and 90 minutes after the nitrate load. Samples of saliva were taken at the same time points, directly after NO measurements.
Nitrate and nitrite in tracheal and oral EBC in tracheostomised patients
Thirteen tracheostomised patients had fasted overnight and, after two baseline collections (−30 and 0 minutes), all subjects ingested sodium nitrate as above and oral and tracheal EBC were collected 60 minutes thereafter.
Orally exhaled and nasal NO, nitrate and nitrite in non-tracheostomised healthy subjects
Thirteen non-tracheostomised healthy subjects had fasted overnight and measurements of nasal NO and oral FENO were made and collection of oral EBC, nasal condensate, and saliva were performed before (−30 and 0 minutes) and 60 minutes after a nitrate load as described above. Nasal NO and oral FENO measurements were made using the Niox system and oral EBC and nasal condensate were collected as described above. At 65 minutes after nitrate ingestion, directly after the 60 minute sampling, the subjects were asked to rinse their mouths with an antibacterial mouthwash solution (25 ml 0.2% chlorhexidine diacetate with 0.01% menthol) for 30 seconds and all measurements were repeated after 5 minutes.
Oral and salivary nitrate and nitrite after different mouthwash procedures
Oral EBC and saliva were collected from 15 healthy subjects before and 5 minutes after mouthwash with dH2O (25 ml) for 30 seconds, and subsequently 5 minutes after mouthwash with chlorhexidine as described above.
Statistical analysis
NO output was calculated as: [NO] (ppb or nl) × V˙ (l/min) = nl/min. This was done to enable comparison between nasal and lower airway NO production. Baseline values for NO output and nitrite and nitrate in EBC and saliva were calculated as the mean of two repeated samples (−30 and 0 minutes). All results are presented as mean (SE). The Mann-Whitney test, the Wilcoxon signed rank test, and the Spearman rank correlation test were used as indicated. If there were more than two repeated measures, a significant Friedman test was a prerequisite before further testing with Wilcoxon. All calculations were made using the Prism software Version 4 (Graphpad Software Inc, San Diego, CA, USA).
RESULTS
Oral v tracheal comparisons in tracheostomised subjects
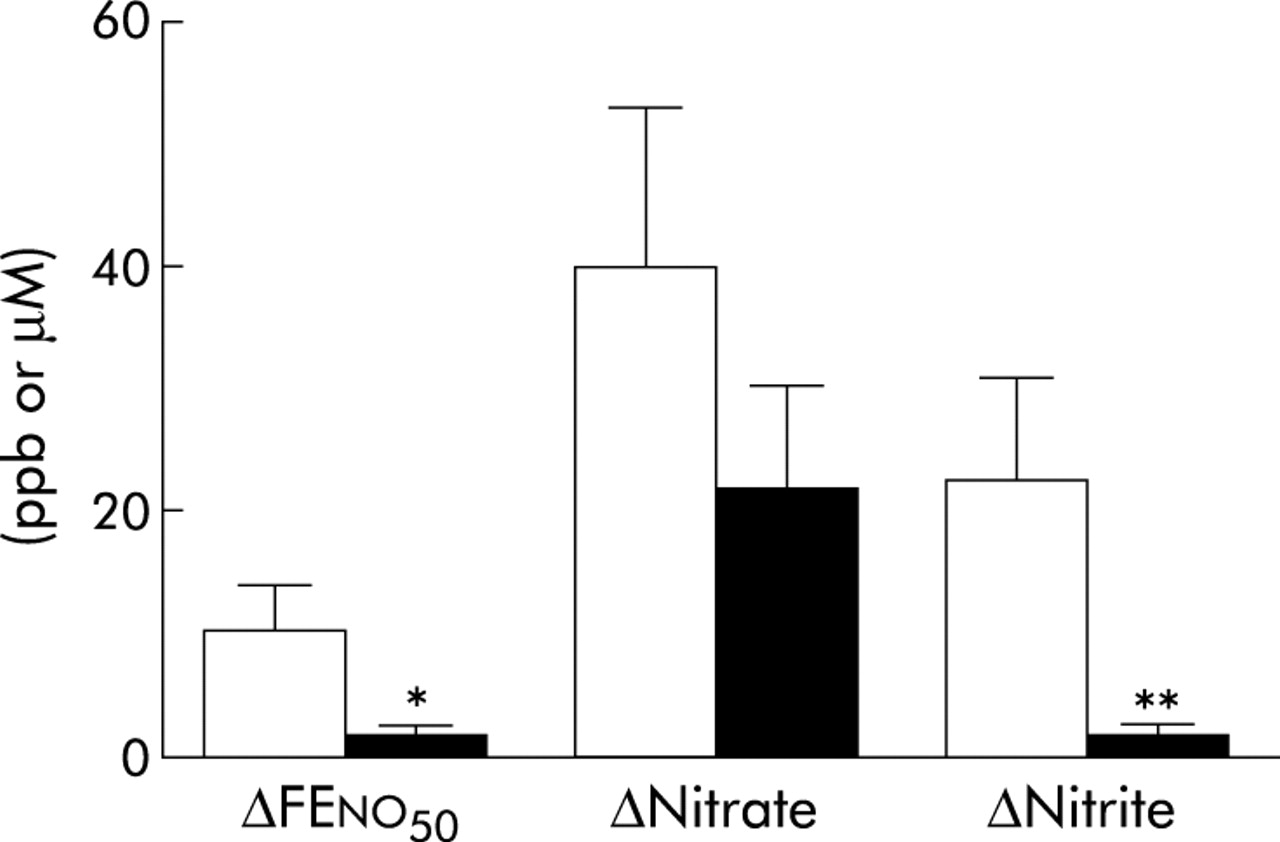
At baseline, mean NO output was approximately twice as high during oral exhalation as during tracheal exhalation (table 2). After nitrate ingestion, oral FENO increased steadily to a maximum at 60 minutes (fig 1A). NO output had then increased by almost 90% compared with baseline levels. At 90 minutes oral FENO had started to return to baseline levels (17 (2.8) ppb at 90 minutes) but was still significantly increased (p<0.05, Wilcoxon test). No significant change in the amount of NO exhaled from the tracheostomy was seen during the total observation period of 90 minutes after nitrate ingestion. The nitrate concentration in the saliva showed a maximal 12-fold increase at 60 minutes (table 3) and the nitrite concentration showed a maximal 18-fold increase at 90 minutes (1167 (289) µM) after nitrate ingestion. The condensate yield was 18 (1) µl/l exhaled air during oral condensate sampling (not different from non-tracheostomised subjects, see below) and 14 (1) µl/l during tracheal sampling (p<0.05, Wilcoxon test). Baseline nitrate concentrations were similar in oral and tracheal EBC, whereas nitrite concentrations were approximately twice as high in oral EBC as in tracheal EBC (table 2). At 60 minutes the nitrate concentration had increased in both oral and tracheal EBC by 284% and 214%, respectively (fig 1C). The absolute increase in nitrate was of a similar magnitude in oral and tracheal EBC (fig 2). There was a 23-fold increase in nitrite concentration in oral EBC 60 minutes after the nitrate load and a fourfold increase in tracheal EBC at the same time point (fig 1B). The absolute increase in FENO was fivefold higher and the increase in EBC nitrite was 12-fold higher during oral exhalation compared with tracheal exhalation (fig 2). Interestingly, salivary levels of nitrate and nitrite as well as nitrite in oral EBC and NO output were all significantly lower in tracheostomised subjects than in non-tracheostomised subjects (table 2). All these differences had disappeared 60 minutes after nitrate ingestion.
Baseline (fasting) values (mean of two consecutive measurements for each subject) for NO output (nl/min) and nitrate and nitrite in condensate and saliva (µM)
Salivary nitrite and nitrate concentrations (µM) before and after nitrate ingestion and 5 minutes after chlorhexidine mouthwash

Effects of nitrate ingestion (10 mg/kg body weight at 0 min) on (A) FENO values, (B) nitrite and (C) nitrate concentrations in oral (filled circles) and tracheal (open circles) exhaled breath condensates in tracheostomised patients (n = 10–13). **p<0.01, ***p<0.001 compared with baseline (0 min; Wilcoxon test).
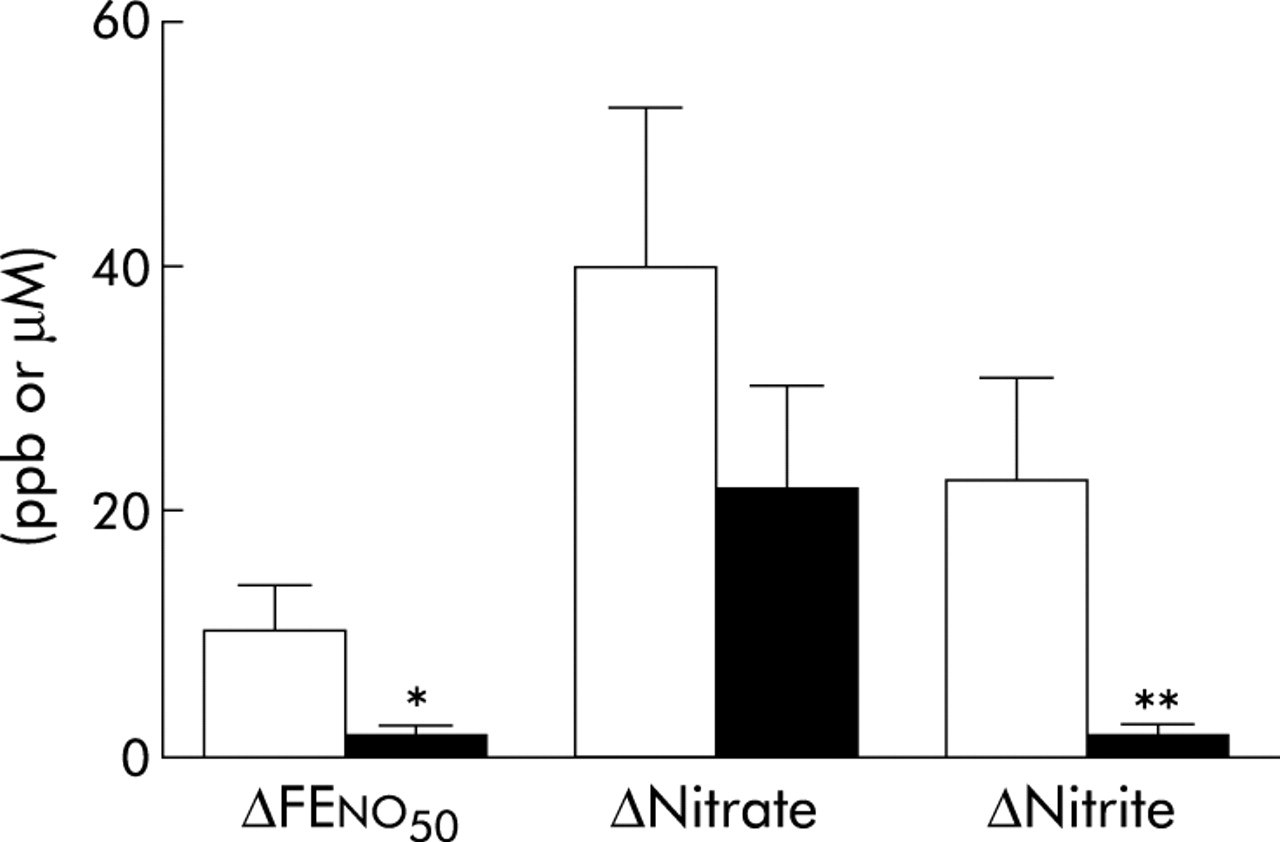
Absolute increase in FENO, nitrate, and nitrite in exhaled breath condensate during oral (open bars) and tracheal (filled bars) sampling 60 minutes after nitrate ingestion (10 mg/kg body weight) in tracheostomised subjects (n = 10–13). *p<0.05, **p<0.01 (Wilcoxon test).
Oral v nasal comparisons in non-tracheostomised subjects
Baseline mean NO output was approximately fivefold higher in the nasal airways than in the lower airways (table 2). Oral FENO increased as expected after the nitrate load (from 17.7 (1.9) ppb at 0 minutes to 26.7 (2.5) ppb at 60 minutes; p<0.001, Wilcoxon). However, nasal NO levels were not affected by nitrate ingestion (table 4). Nitrite in saliva showed an expected ninefold increase 60 minutes after nitrate ingestion (table 3). Nasal condensate yield was 26 (1) µl/l air which was significantly higher than for oral EBC (18 (1) µl/l; p<0.01, Wilcoxon test). Baseline levels of nitrate were similar in nasal condensate and oral EBC, whereas the mean nitrite concentration was 60% higher in oral EBC than in nasal condensate (table 2). After nitrate ingestion there was a fivefold increase in nitrite concentration in oral EBC at 60 minutes (from 3.5 (0.7) µM at 0 minutes to 17.8 (3.4) µM at 60 minutes; p<0.001, Wilcoxon test), similar to findings in the tracheostomised subjects (see above). However, nitrate and nitrite concentrations in nasal condensate did not change after the nitrate load (table 4). The increase in FENO and the increase in nitrite concentration in oral EBC both correlated with the increase in salivary nitrite levels (table 5).
Nasal NO levels (ppb) and nitrite and nitrate in nitrate condensate (µM) before and after nitrate ingestion and 5 minutes after chlorhexidine mouthwash
Correlation coefficients for relations between changes in salivary nitrite versus changes in nitrite in oral EBC and FENO after nitrate ingestion and after chlorhexidine mouthwash
Evaluation of antibacterial mouthwash
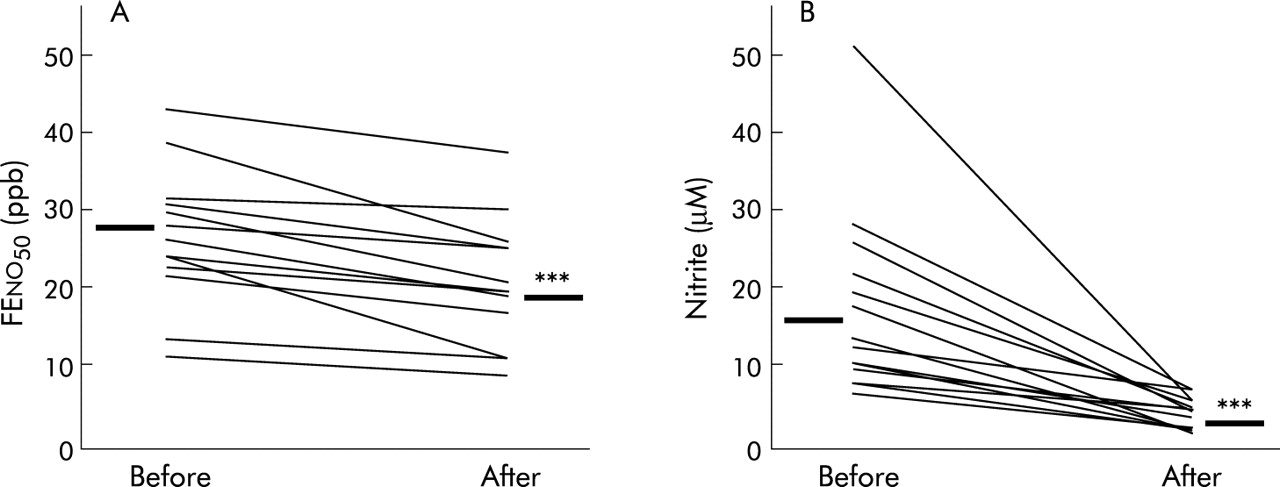
Mouthwash with deionised water did not change the levels of nitrate or nitrite in saliva or oral EBC (table 6). In contrast, chlorhexidine mouthwash significantly reduced nitrite levels in saliva and oral EBC while nitrate levels remained unaffected. Sixty minutes after a nitrate load, antibacterial mouthwash reduced oral FENO in all subjects (mean reduction 20%; fig 3A) but it was still higher than baseline, whereas nitrite levels in oral EBC were restored to baseline levels by the mouthwash (mean reduction 81%; fig 3B). Nitrate levels in EBC were not affected by chlorhexidine (not shown). The reduction in FENO and nitrite levels in oral EBC by chlorhexidine both correlated with the reduction in salivary nitrite levels (table 5). Amylase activity was not detected in any of the condensate samples from either tracheostomised or non-tracheostomised subjects, whereas high amylase activity was noted in saliva samples (>5000 IU/l).
Levels of nitrite and nitrate (μM) in oral EBC and saliva before and after mouthwash with deionised water and chlorhexidine
{kind=link}
{kind=link}
{kind=link}
Effect of antibacterial mouthwash (chlorhexidine 0.2%) given 60 minutes after nitrate ingestion on (A) oral FENO values and (B) oral EBC nitrite levels in non-tracheostomised subjects (n = 13). ***p<0.001 v before mouthwash (Wilcoxon test).
DISCUSSION
We have confirmed that orally exhaled NO is only partly derived from the airways below the upper trachea in respiratory healthy subjects, and that there is a significant contribution of NO from the oropharynx formed via the reduction of nitrite in saliva. We have been able to extend the knowledge on nitrite derived NO formation in the airways by sampling from the lower or nasal airways, isolated from the oropharyngeal tract, before and after ingestion of nitrate. Furthermore, we have studied the possible oropharyngeal contribution of nitrite and nitrate in oral EBC using a standard technique.
After ingestion, nitrate is rapidly absorbed in the gastrointestinal tract and the plasma concentration increases.10,21 Nitrate is then actively taken up by the salivary glands in a plasma concentration dependent way and salivary nitrate concentrations increase also.10 In the oral cavity facultative anaerobic bacteria in the deep crypts of the base of the tongue reduce nitrate to nitrite in the course of respiration and, thus, salivary nitrite levels are also increased.10–12 Some of this nitrite is reduced to NO in the oral cavity which contributes to levels of NO measured in exhaled breath.11,21 Our data indicate that nitrate is also taken up directly from the circulation by the lower airways since nitrate concentrations were increased in the tracheal EBC of our tracheostomised subjects 1 hour after nitrate ingestion. The increase in the lower airways may be explained by passive diffusion from plasma. This would be in line with previous studies in which the distribution volume for nitrate was found to be large (approximately 30% of body weight).22,23
Experiments with radioactively labelled nitrate have shown that a substantial part of the ingested radioactive nitrogen cannot be recovered in faeces or urine.23 One explanation is that the lost nitrate undergoes metabolism to gaseous products and is exhaled in the breath as, for example, NO.11 We show here that some of the ingested nitrate is excreted in the respiratory tract also by aerosolisation of airway epithelial lining fluid and saliva. Interestingly, the airway mucosal uptake of nitrate from plasma seems to be restricted to the lower airways since nitrate levels in nasal condensate were not affected by a nitrate load.
We did not see a change in nitrite concentration in nasal condensate after nitrate ingestion, which indicates that we did not have any contamination from the pharynx during nasal condensate collection with our method. Others have collected breath condensate from the nose during normal tidal breathing24 but, if such a method is used, the origin of the collected fluid will be a mixture of the lower airways, pharynx, and the nasal airways.
Although nitrite levels in nasal condensate were unchanged, we found a significant increase in the nitrite concentration in tracheal condensate after the nitrate load. This increase may not rely on passive diffusion from the circulation since the plasma nitrite concentration is usually very low and is not readily increased to high concentrations by nitrate ingestion. It is well known that nitrite is rapidly oxidised to nitrate in the presence of oxyhaemoglobin. Instead, the increase in lower airway nitrite may be due to xanthine oxidase activity in the pulmonary epithelium25,26 or bacterial activity in the trachea of tracheostomised subjects. Patients with a permanent tracheal cannula are often colonised by bacteria27 such as Pseudomonas aeruginosa, both in the trachea and in biofilm formation on the cannula tube.28P aeruginosa is capable of reducing nitrate to nitrite both under anaerobic and aerobic conditions.29 Thus, bacterial activity could be a reason for the increased nitrite concentration seen in tracheal EBC in our tracheostomised subjects. However, no relation between the presence of P aeruginosa (or any of the other bacteria detected in our sample of tracheostomised subjects) and the level of nitrite or nitrate in tracheal EBC could be found. Interestingly, the tracheostomised subjects had lower baseline levels of nitrite and nitrate in saliva, nitrite in oral EBC, and exhaled NO than non-tracheostomised subjects. These differences could be due to the lower nitrate content in saliva, indicating an altered saliva production or nitrate uptake in the tracheostomised subjects or, alternatively, an altered bacterial activity in the oropharyngeal tract. If this is due to the tracheostomy, the higher age in this group or other causes is not known.
In this study we have shown that changes in salivary nitrite levels correlate with changes in exhaled NO and nitrite in oral EBC, both after nitrate ingestion and chlorhexidine mouthwash. Thus, nitrite in saliva is an important contaminant for EBC collections as well as for exhaled NO measurements.11,21 Indeed, the maximal oropharyngeal influence on EBC nitrite levels is of much greater magnitude than that on exhaled NO. A reduction of nitrite in saliva and oral EBC was seen after mouthwash with chlorhexidine but not with deionised water, similar to earlier findings for exhaled NO.11 Nitrate levels were not affected by chlorhexidine which is a potent antibacterial agent with well documented effects in the oral cavity.30 Together these data clearly indicate that nitrite formation in the saliva—and thus the levels found in oral EBC—are dependent on bacterial activity in the oropharyngeal tract. Several groups have reported that the nitrite and/or nitrate concentration in EBC is increased in airway diseases, particularly asthma and CF,16,17,31–34 and the general postulate has been that the measured levels of nitrite and nitrate are oxidation products of NO. Our results show that both nitrite and nitrate levels in the lower airways can also increase after nitrate ingestion, and an antibacterial mouthwash reduces the presence of nitrite in EBC. Thus, nitrite in EBC may primarily be a marker of oropharyngeal bacterial activity and not a marker of NO formation and airway inflammation. However, in pneumonia induced acute lung injury, nitrite formation also seems to increase in the lower airways below the tracheal tube,35 but these conditions are rarely connected with increases in exhaled NO indicating that this nitrite is also of bacterial origin. After cigarette smoking (tobacco smoke contains high amounts of NO) only nitrate and not nitrite seems to be increased in EBC.36,37 Although these studies indicate that nitrate in EBC may be related to the presence of NO, further studies are needed to elucidate the relation between the formation of NO, nitrite, and nitrate in the lower airways in health and disease where nitrite and nitrate should be analysed and reported separately.
It has been suggested that NO formation from nitrite in the lower airways could contribute to exhaled NO levels in acute inflammation when the pH in the airways is reduced.38 However, we found substantial amounts of nitrite derived NO formation from the saliva where the pH is usually close to 7 (between 6 and 8), while no such NO formation could be detected in the lower airways below the upper trachea. The reduction of nitrite to NO may also be enhanced by the presence of reducing agents—for example, ascorbate11,39—and it is thus suggested that the presence of such agents may be the dominating factor controlling the reduction of nitrite at a neutral pH. Some bacteria are also able to reduce nitrite to NO.29 However, commensal bacteria very rarely use this denitrifying pathway but instead use a short cut resulting in the formation of ammonia.40
We have shown that the oropharyngeal tract contributes substantially to nitrite and, to some extent, nitrate in EBC collected in a standardised way, and the condensate yield was higher during oral than tracheal normal tidal breathing. This was seen in spite of the fact that amylase activity was not detected in the samples which, in turn, shows that measurements of amylase activity are not adequate to rule out contamination from the oropharyngeal tract. Our findings indicate that a significant portion of respiratory droplets15 that “survive” all the way beyond the lips during exhalation are formed in the very proximal airways including the oropharyngeal tract. Respiratory droplets are most likely formed also in the peripheral airways,41 but it is not known what proportion of these peripheral droplets actually travel all the way to the exterior during exhalation without being deposited more centrally in the airways.
We conclude that plasma nitrate is taken up by the salivary glands and the lower airways but not by the nasal airways. Nitrite formation from nitrate in the human respiratory tract is confined to the oropharyngeal tract and trachea and probably involves bacterial activity. The further reduction of nitrite to NO is restricted to the oropharynx, at least in our subjects without inflammatory airway disease. Another significant finding is that a major part of the nitrite found in EBC originates in the oropharyngeal tract above the upper trachea, indicating proximal generation of respiratory droplets. Nitrate in EBC seems to originate both in the oropharynx and the lower airways but is influenced by plasma levels and thus dietary intake in the form of, for example, green leaved vegetables.
Acknowledgments
The authors gratefully acknowledge Ms Margareta Stensdotter and Mrs Carina Nihlén for their expert technical assistance during the biochemical analyses, and are grateful to Dr Kenth Johansson, Institute for Surface Chemistry, Stockholm, for help with plasma treatment of condenser tubes.
REFERENCES
Footnotes
-
This study was supported by the Swedish Research Council, Medicine (no 10354), the Swedish Heart-Lung Foundation, the Swedish Asthma and Allergy Association’s Research Foundation, the Swedish Knowledge Foundation, Karolinska Institutet and Aerocrine AB.
-
Competing interests: HM, DCT and US declare no competing interests. KA and EW are co-founders and shareholders of Aerocrine AB. KA has received research funding from Aerocrine. EW is a member of the board of Aerocrine.