Article Text

Abstract
Background: Non-specific bronchial hyperresponsiveness (NSBH) is a known predictor of accelerated rate of decline in lung function in smokers. Polymorphisms of the β2 adrenergic receptor (ADRB2) have previously been associated with NSBH and bronchodilator response (BDR) in asthmatics. Based on these associations, we hypothesised that ADRB2 polymorphisms would be associated with NSBH and BDR as well as an accelerated rate of decline in lung function among smokers.
Methods: The prevalence of two ADRB2 polymorphisms, Arg16→Gly and Gln27→Glu, was examined in 587 smokers chosen from the NHLBI Lung Health Study for having the fastest (n=282) and slowest (n=305) 5 year rate of decline in forced expiratory volume in 1 second (FEV1; mean ΔFEV1 −4.14 and +1.08% predicted/year, respectively).
Results: Contrary to our hypothesis, no ADRB2 allele or haplotype was associated with NSBH, BDR, or rate of decline in lung function. However, there was a significant negative association between heterozygosity at position 27 and a fast decline in lung function (adjusted odds ratio 0.56, 95% CI 0.40 to 0.78, p=0.0007).
Conclusions: Heterozygosity at position 27 may be protective against an accelerated rate of decline in lung function. The polymorphism at position 16 does not contribute to the rate of decline in lung function, measures of NSBH, or BDR in smokers.
- bronchial provocation tests
- lung volume measurements
- chronic obstructive pulmonary disease
- genetics
- β2 adrenergic receptors
Statistics from Altmetric.com
- bronchial provocation tests
- lung volume measurements
- chronic obstructive pulmonary disease
- genetics
- β2 adrenergic receptors
Non-specific bronchial hyperresponsiveness (NSBH) is a known risk factor for the respiratory symptoms and disease progression in chronic obstructive pulmonary disease (COPD). In cross sectional studies, smokers and non-smokers who have NSBH have lower baseline forced expiratory volume in 1 second (FEV1)1 and higher respiratory symptom scores2 than those who do not have NSBH. Furthermore, smokers and non-smokers with NSBH have been shown in longitudinal studies to have a faster rate of decline in FEV1.3,4
Although cigarette smoking contributes to both NSBH and respiratory symptoms such as chronic cough, dyspnoea and wheezing, the association between NSBH and respiratory symptoms exists independently of cigarette smoking.2 NSBH is also closely associated with atopy5 and is a diagnostic feature of asthma.6 The observation that NSBH is associated with both asthma and COPD supports the longstanding hypothesis that a common determinant underlies both these conditions.7
Polymorphisms in the β2 adrenergic receptor (ADRB2) have previously been shown to be associated with asthma severity,8,9 NSBH,10,11 bronchodilator response,12–15 and level of lung function.16 Wang et al showed that the ADRB2 Arg16 allele was a risk factor for asthma and there was a significant interaction with cigarette smoking.17 However, Ho et al found that Arg16 was less prevalent in Chinese patients with COPD and that the Gln27 allele was associated with more severe airflow obstruction.18 The Arg16→Gly and Gln27→Glu polymorphisms in ADRB2 are known to affect agonist induced receptor downregulation in vitro19,20 and in vivo.21 We hypothesised that, if these polymorphisms affect NSBH in asthma, they may also contribute to NSBH in COPD, thus potentially modifying bronchodilator response (BDR) and rate of decline in lung function (ΔFEV1/year).
Associations of NSBH with FEV1, rate of decline in lung function, and respiratory symptoms of COPD have been shown in the NHLBI Lung Health Study (LHS) cohort.22 The LHS (phase I) was a multicentre randomised clinical trial that measured lung function in 5887 subjects for 5 years.23 Measurements of NSBH to methacholine and BDR were made in all subjects at baseline. From the continuing smokers in the LHS I cohort we selected the 282 with the fastest rate of decline in FEV1 over 5 years and the 305 with the greatest increase in FEV1 over the same time period. The prevalence of the Arg16→Gly and Gln27→Glu polymorphisms in the ADRB2 was compared in these groups using established polymerase chain reaction (PCR) methods. We also tested whether there was a relationship between these polymorphisms and NSBH measured using inhalation of methacholine and response to an inhaled β adrenergic agent (isoproterenol). This is the first study to examine the relevance of these polymorphisms to NSBH, BDR, and rate of decline in lung function in smokers.
METHODS
Study subjects
Subjects were selected from participants in phase I of the Lung Health Study. The design of this multicentre randomised clinical trial has been described more extensively elsewhere.23 Study participants were current smokers aged 35–60 with mild to moderate airflow obstruction (FEV1 55–90% predicted and FEV1/FVC ⩽0.70). Exclusion criteria included serious illnesses such as cancer, heart attack or stroke, or other important conditions that required medical treatment. The primary outcome variable was rate of decline in FEV1 over a follow up period of 5 years. Of the 3216 continuing smokers in this cohort, 282 were chosen for their rapid decline in lung function (decline in % predicted FEV1 >3.0% per year), and 305 were selected with no decline in lung function over the same period (increase in % predicted FEV1 >0.4% per year). All selected participants were white.
Genotyping methods
Genotyping of the polymorphisms at positions 16 and 27 was performed by restriction enzyme digestion of PCR products as previously described.9 Briefly, 12 μl of a 50 μl PCR product was digested with NcoI at an introduced restriction site to determine the genotype encoding amino acid 16. In addition, 12 μl of the same PCR product was digested with BbvI to identify the presence or absence of a naturally occurring restriction site at the locus encoding amino acid 27. The products of each PCR reaction were resolved by electrophoresis on 3% agarose gels stained with ethidium bromide. Template-free controls and known genotype controls were included in each experiment. Genotypes were scored without knowledge of the phenotypes by two independent observers. The samples were re-genotyped if there was any disagreement concerning the genotype. Twenty samples were selected at random and were sequenced to confirm the genotyping protocol. All samples showed the same genotypes for both polymorphisms as with the restriction enzyme analysis.
Statistical analysis
The Gly16 and Gln27 allele frequencies and allelic associations with rate of decline in lung function were analysed using the χ2 test for 2×2 contingency tables. The associations were also analysed by multiple logistic regression to adjust for potential confounding factors. The outcome variables of interest were BDR, NSBH, and a dichotomous variable related to rate of decline in lung function—that is, rapid decline or no decline. Potential confounding factors included in the analysis were smoking history (pack years), age, sex, and initial level of lung function (pre-bronchodilator FEV1 % predicted). Two factor interactions were also investigated in the models. NSBH was expressed as a two point methacholine dose-response slope as previously described.24 Mean BDR in the different genotypic groups was compared by ANOVA. Differences in mean NSBH were compared by the Wilcoxon test since these data were not normally distributed. All tests were performed using the JMP Statistics software package (SAS Institute Inc).
Although we confined our analyses to the white individuals in the study, there could be genetically distinct subgroups in this population because the LHS was a multicentre study. To minimise the possibility of a spurious association due to population stratification, we therefore added “centre” as a variable in the regression analysis to test for any significant association.
Haplotype frequencies were estimated using the expectation maximisation (EM) algorithm25 since haplotypes could not be discerned directly from double heterozygotes. The Arlequin software package was used to perform these estimations, to test for linkage disequilibrium, and for Hardy-Weinberg equilibrium.26
RESULTS
The characteristics of the study subjects are summarised in table 1. For both polymorphisms the overall observed distribution of homozygotes and heterozygotes conformed to expectations based on Hardy-Weinberg analysis. There was strong linkage disequilibrium between alleles at the two loci (D′=100%, p<0.000001) as shown by other investigators.9,11,12 Power analyses showed that our study design had sufficient power to detect a 1.3 fold difference in BDR and a 1.4 fold difference in methacholine NSBH between genotypic groups for the Arg16→Gly polymorphism, and a relative risk for rapid decline in lung function as low as 1.6. There was similar power to detect associations with the Gln27→Glu polymorphism (data not shown).
Characteristics of the 587 study subjects
Contrary to our hypothesis, no allele, genotype or haplotype was associated with methacholine NSBH or with BDR (table 2). The allele frequency for Glu27 was similar in subjects with a fast decline in lung function (0.47) and in those with no decline in lung function (0.48), and the rate of decline in lung function was not associated with the estimated haplotypes (table 3).
Analysis of bronchodilator response and methacholine responsiveness stratified by β2 adrenergic receptor genotype
Estimated haplotype frequencies among smokers with a fast decline in lung function and in those with no decline in lung function
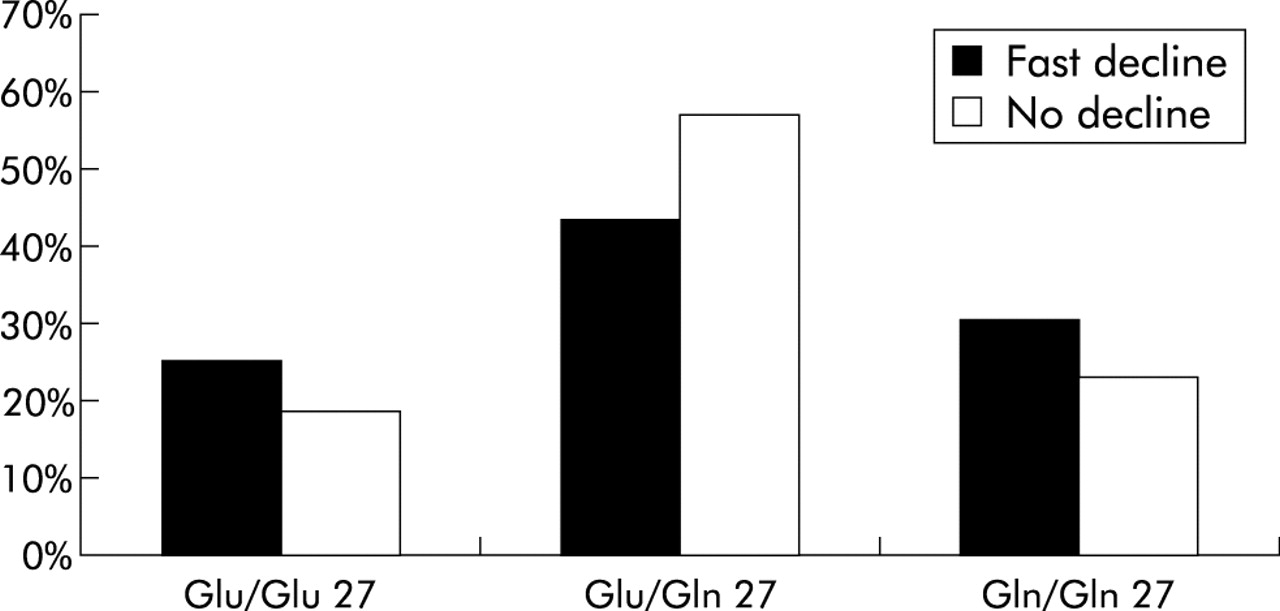
However, the Glu27/Gln27 heterozygous genotype was significantly less prevalent in smokers with a rapid rate of decline in lung function (odds ratio 0.57, 95% CI 0.41 to 0.80, p=0.0009, table 4). Figures 1 and 2 show the distribution of genotypes within the two groups of smokers. The excess heterozygotes in those with no decline in lung function resulted in a deviation from Hardy-Weinberg equilibrium (p=0.03) in this group. There was a similar trend away from Hardy-Weinberg equilibrium in the subjects in whom there was a fast decline in lung function, but this did not reach statistical significance (p=0.11). However, when the two groups were combined there was a good fit with Hardy-Weinberg equilibrium (p=0.91).
Distribution of Glu27/Gln27 genotypes among smokers with a fast decline in lung function and in those with no decline in lung function

Genotype distribution (position 16) in subjects with a fast decline in lung function and in those with no decline. There was no significant difference in genotype frequencies between the groups (p=0.46 for a 3×2 contingency table).

{kind=link}
{kind=link}
Genotype distribution (position 27) in subjects with a fast decline in lung function and in those with no decline. A significant difference in genotype frequency was seen between the two groups (p=0.004 for a 3×2 contingency table). Heterozygosity for the Glu/Gln27 allele was significantly more frequent in those with no decline in lung function, suggesting a protective effect of this genotype.
These data suggest that the Glu27/Gln27 genotype has a protective role against a rapid decline in lung function. However, it is important to ensure that this association was not due to chance effects of β2 adrenergic receptor status on smoking exposure, so a one way ANOVA was performed to show that there was no significant difference (p=0.98) in smoking history between heterozygotes and homozygotes for the Glu27/Gln27 polymorphism. To adjust this result for potentially confounding factors a logistic regression was also performed (table 5); the association between heterozygosity for the polymorphism at position 27 and the rate of decline remained significant in this analysis. The adjusted odds ratio for the Glu27/Gln27 genotype and the rate of decline in lung function was 0.56 (95% CI 0.40 to 0.78, p=0.0007). When “centre” was added as a variable in the regression analysis, the association between Glu27/Gln27 heterozygosity and rate of decline in lung function was still significant (p=0.0018).
Multiple logistic regression for fast decliners versus no decliners
Since ADRB2 polymorphisms have previously been associated with NSBH and BDR, we tested for possible interactions between these phenotypes and the Glu27/Gln27 genotype. There was a significant interaction between heterozygosity at position 27 and NSBH to methacholine on the rate of decline (p<0.0001), but there was no interaction between Glu27/Gln27 genotype and BDR (p=0.41).
DISCUSSION
ADRB2 polymorphisms occur commonly in the human population and have been associated with a number of asthma phenotypes.8,9,11,12,27–29 The aim of this study was to investigate the association between ADRB2 polymorphisms and BDR, NSBH, and the rate of decline in lung function in smokers. In contrast to results in patients with asthma, we found no association between ADRB2 polymorphisms and either BDR or NSBH. However, there was a significantly lower prevalence of heterozygous Glu27/Gln27 individuals among the smokers with a fast decline in lung function, suggesting a protective effect of this genotype.
Previous association studies of ADRB2 polymorphisms and specific phenotypes related to airway disease and function have been reviewed by Joos et al.30 Despite considerable controversy and conflicting results, the most consistent finding was a relationship between the polymorphism at position 16 and BDR. However, we found no association between BDR and any allele, genotype, or haplotype in this study. This could be related to the sampling strategy used. Because we were primarily interested in polymorphisms which affect the rate of decline in lung function, we selected those subjects with the most rapid and the slowest decline in lung function rather than a random sample of subjects. Despite this, the distribution of BDR as measured by the % increase in FEV1 was normally distributed in the group as a whole. However, different factors might influence BDR in subjects with a rapid decline in lung function compared with those with a slow decline in lung function.
In subjects with asthma NSBH has been associated with specific ADRB2 polymorphisms; specifically, asthmatics who had the Gly16/Gln27 haplotype were more responsive to methacholine than subjects with other haplotypes.15 We found no association of any allele, genotype or haplotype with airway responsiveness in our group despite a wide range of responsiveness. Because methacholine responsiveness is a powerful predictor of the rate of decline in lung function in smokers, the distribution of methacholine responsiveness (expressed as slope) was not normally distributed in our subject sample and, in fact, showed a bimodal distribution. Again this distribution, which was related to subject selection, could have masked a real association between ADRB2 polymorphisms and NSBH. Despite the lack of an association between any polymorphism and methacholine responsiveness, we found a strong interaction between the ADRB2 genotype, methacholine responsiveness, and rate of decline in lung function. Heterozygosity for the Gln27→Glu polymorphism was only protective against a rapid rate of decline in lung function in individuals who had low responsiveness to methacholine (values less negative than −16.5). These individuals represented 72.4% of the total study participants. The individuals with more responsive airways may have been exposed to other risk factors for NSBH (either environmental of genetic factors) that overwhelmed the protective effect of a Gln27/Glu genotype.
The finding that the heterozygous state displays a different phenotype from either homozygous state is unusual but is a well recognised phenomenon termed “heterozygous advantage”. Several studies in a variety of diseases have shown a distinct phenotype in subjects with heterozygosity at specific loci.31–34 For example, a study of the role of the secretor blood group in psoriasis showed that heterozygosity had a strong protective effect (odds ratio 0.17) even though the allele frequencies did not differ between patients and controls.35
Although one possible explanation for the apparent association could be a type 1 error, the strength of the association and the strongly positive interaction with methacholine responsiveness prompted us to speculate about biologically plausible explanations for the finding. One possible mechanism for a heterozygote advantage is an interaction between the wild type and mutant protein isoforms in heterozygous cells. This is a plausible mechanism for receptor molecules, especially if they form dimers. A “heterodimeric” receptor consisting of a wild type and a mutant protein may have different agonist binding, signal transduction, or agonist promoted desensitisation properties than wild type or mutant homodimers. Several lines of evidence indicate that ADRB2, like other G protein coupled receptors, form dimers. The formation of ADRB2 dimers was shown to have functional effects on receptor stimulated adenylate cyclase activity.36 In addition, ADRB2 ligands have been shown to promote receptor dimerisation.36,37 The results of a study of a mutant ADRB2 (Cys341Gly) suggested that the dimer is the active form of the receptor.38
If there is a heterozygote advantage, a possible mechanism could be that Gln/Glu “heterodimers” have altered functional characteristics compared with either Gln/Gln or Glu/Glu homodimers. The Gln27→Glu polymorphism does not occur in the sixth transmembrane region that is important in dimer formation.36 However, a synthetic peptide derived from this region resulted in only a 69% reduction in dimer formation indicating that other regions may also influence dimerisation.
An alternative explanation for the excess of heterozygotes among those with a slow decline in lung function could be related to linkage disequilibrium between the Gln27→Glu polymorphism and another polymorphism within the ADRB2 gene or the immediate region. The ADRB2 gene contains a small open reading frame (ORF) upstream to the receptor transcription start point. This ORF encodes a 19 amino acid peptide that modulates receptor translation.39 A polymorphism in this peptide (Arg19→Cys) was shown to affect ADRB2 expression at the protein level.40 In human airway smooth muscle cells ADRB2 receptor density was approximately twofold higher in cells with the Cys isoform than in those with the Arg isoform.40 However, because of the strong linkage disequilibrium between the two polymorphisms, Glu27 (which is associated with protection against agonist induced receptor downregulation) is almost always found with Arg19.40,41 In a recent study Glu27 was found exclusively with Arg19 and vice versa in all 396 haplotypes delineated.41 Likewise, Gln27 was found exclusively with Cys19. The only individuals who have both alleles associated with high ADRB2 receptor density (Glu27 and Cys19) are therefore those that are heterozygous at both loci. The heterozygous state at position 27 is thus a marker for the compound genotype. Individuals with this genotype would have one allele conferring high expression at baseline (Cys19) and one allele (Glu27) that confers resistance to downregulation after agonist exposure. The overall physiological effect of this combination may confer greater protection from loss of lung function than either homozygotes for Cys19 (who will also be homozygous for Gln27) or homozygotes for Glu27 (who will also be homozygous for Arg19).
Another possible explanation for these data is a spurious association because of population stratification. Although we confined our analyses to the white individuals in the study, there could be genetically distinct subgroups in this population because the LHS was a multicentre study. The fact that the two polymorphisms were in Hardy-Weinberg equilibrium and that there was no difference in allele frequencies between subjects with a fast decline in lung function and those in whom lung function did not decline provides a powerful argument that the association of this polymorphism with the rate of decline was not confounded by population structure. We also added “centre” as a variable in the regression analysis and showed that the association of Glu27/Gln27 heterozygosity with rate of decline in lung function was still significant in this analysis.
In addition to analysing individual polymorphisms, we also estimated haplotype frequencies. Since no association was found with the haplotypes, there may be concern regarding the accuracy of the estimates since these were not measured directly but were estimated using the EM algorithm. However, the results of a recent study have shown that estimations of haplotype frequency based on the EM algorithm accurately reflect the true frequencies, particularly for common polymorphisms such as those studied here.42 There are many additional polymorphisms in the ADRB2 gene and these could be used further to define the haplotypes in our population. However, Drysdale et al performed a comprehensive survey of ADRB2 haplotypes and showed that the Arg16/Gly16 and Glu27/Gln27 polymorphisms are sufficient to distinguish the three common haplotypes in white subjects.41 Another seven haplotypes exist in the white population but they are all rare and the combined frequency of these haplotypes is only 5.5%. It is therefore unlikely that additional haplotypic data would substantially change our results.
In summary, this is the first study to address the possible influence of the ADRB2 polymorphisms on BDR and methacholine NSBH as well as the rate of decline in FEV1. The Arg16→Gly polymorphism at position 16 was not associated with the rate of decline in lung function, measures of NSBH, or BDR in smokers. However, there was a novel negative association between heterozygosity at position 27 and a fast decline in lung function, despite the lack of an association with NSBH or BDR. Possible explanations for this observation are molecular interaction of isoform heterodimers or linkage disequilibrium with a second protective allele.
Acknowledgments
This study was supported by a grant from the Canadian Institutes of Health Research. The Lung Health Study was supported by contract N01-HR-46002 from the Division of Lung Diseases of the National Heart, Lung, and Blood Institute. Dr Joos was supported the Swiss National Science Foundation, Novartis Research Foundation, Uarda-Frutiger Foundation and the Swiss Society of Pneumology. Dr Sandford was supported by a Parker B Francis Fellowship and is the recipient of a Canada Research Chair in genetics. The authors would like to thank Helen Voelker for expert assistance with the statistical analysis.