Article Text

Abstract
Background: The extracellular matrix metalloproteases (MMPs) produced by tumour and stromal cells are believed to play a key role in tumour cell invasion and metastasis. Malignant mesothelioma is a highly aggressive tumour. Previous studies have shown that malignant mesothelioma cells express MMP-1, -2, -3, -7 and -9. However, the regulation of MMP expression in malignant mesothelioma cells has not been thoroughly investigated.
Methods: The effects of a number of growth factors on the secretion of MMP-2, -3 and -9 in malignant mesothelioma cells were studied by substrate zymography. The expression of relevant growth factor receptors in malignant mesothelioma cells was also examined using RT-PCR.
Results: The exposure of malignant mesothelioma cells to different growth factors including epidermal growth factor (EGF), transforming growth factor-α, amphiregulin, heparin binding EGF, β-cellulin (BTC), stem cell factor, insulin-like growth factors I and II, acidic and basic fibroblast growth factors, and hepatocyte growth factor increased secretion of MMP-9 and/or MMP-3. EGF receptor ligands BTC and EGF had the most potent effect on MMP-9 and MMP-3 production, respectively. Production of MMP-2 was not affected by any growth factors used in this study. We have also demonstrated mRNA expression of different growth factor receptors in malignant mesothelioma cells, and found that the tyrosine kinase inhibitor genistein inhibited the increased production of MMPs resulting from stimulation with different growth factors.
Conclusion: This study shows, for the first time, the broad extent of regulation of MMPs by various growth factors in malignant mesothelioma cells.
- matrix metalloproteases
- malignant mesothelioma
- growth factors
Statistics from Altmetric.com
Malignant mesothelioma is an asbestos associated tumour arising from mesothelial lined surfaces, most often the pleural cavities.1 Malignant mesothelioma is a highly aggressive tumour that has a tendency to grow rapidly and invade locally. Necropsic examination has shown metastatic spread in 50% of patients.2 Epidemiological evidence suggests that the number of people dying from mesothelioma each year will increase in Western Europe over the next 20 years.3 Since malignant mesothelioma is resistant to all conventional anticancer treatments, understanding the mechanisms involved in the invasiveness of malignant mesothelioma may provide insights into new therapeutic strategies.
Proteolytic degradation of extracellular matrix and basement membranes is a key component of the process of tumour cell invasion and metastasis. Matrix metalloproteases are a growing family of zinc dependent endopeptidases that currently comprise at least 20 members and are capable of degrading all components of extracellular matrix.4 These enzymes are involved in a variety of physiological and pathological processes including embryogenesis, angiogenesis, tumour invasion, and arthritis.4 Several metalloproteases including MMP-1 (collagenases), MMP-2, MMP-9 (gelatinases), MMP-3, and MMP-10 (stromelysins) have been implicated in tumour cell invasion.5 Previous studies have shown that malignant mesothelioma cells express several members of the MMP family with different substrate specificities.6,7
MMP expression is regulated at both pre and post transcriptional levels. A number of extracellular factors—including growth factors, cytokines and cell contact with extracellular matrix—have been implicated in the regulation of MMP expression in different types of tumour cells.6,8 However, the regulation of MMP activity in malignant mesothelioma cells has not been thoroughly investigated.
We have investigated the effects of different growth factors on production of MMP-2, MMP-3 and MMP-9 in malignant mesothelioma cells. Furthermore, to identify potential molecular mechanisms responsible for regulation of MMPs by various growth factors, we have examined the expression of those growth factor receptors in malignant mesothelioma cells. We have also studied the inhibitory effects of the tyrosine kinase inhibitor genistein on the increased production of MMPs promoted by different growth factors in malignant mesothelioma cells.
METHODS
Reagents
RPMI 1640, fetal calf serum (FCS), penicillin, streptomycin, and l-glutamine were obtained from Life Technologies (Paisley, UK). Gelatin was purchased from SIGMA (St Louis, MO, USA). Fibronectin was purified as previously described.9
Growth factors and tyrosine kinase inhibitor
Epidermal growth factor (EGF), transforming growth factor alpha (TGF-α), amphiregulin (AR), heparin binding EGF-like growth factor (HB-EGF), β-cellulin (BTC), stem cell factor (SCF), insulin-like growth factor (IGF)-I, IGF-II, acidic and basic fibroblast growth factor (aFGF, bFGF), platelet derived growth factor (PDGF)-BB, and hepatocyte growth factor (HGF) were purchased from R & D Systems (Abingdon, UK). The protein tyrosine kinase inhibitor genistein was purchased from Life Technologies (Paisley, UK).
Cells and culture conditions
Six human malignant mesothelioma cell lines were used: STAV-FCS,10 ZL5, ZL34, SPC212, M28K, and M38K. The human fibrosarcoma cell line HT-1080 was purchased from American Type Cells Collection (ATCC). All malignant mesothelioma cell lines were cultured in RPMI 1640 supplemented with 5% FCS, penicillin (100 U/ml), streptomycin (100 μg/ml), and 2 mM l-glutamine. HT-1080 was cultured in RPMI 1640 supplemented with 10% FCS. The cells were maintained in a humidified atmosphere of 5% CO2 in air at 37°C.
RNA isolation, cDNA synthesis, primers and RT-PCR
Total RNA was isolated from cell lines using the guanidinium thiocyanate method. Using a cDNA synthesis kit according to the manufacturer’s instructions (Pharmacia Biotech, Uppsala, Sweden), first strand cDNA synthesis with Not-(T) 18 primers was performed. For cDNA synthesis 5 μg of total RNA were taken. The reverse transcript (1 μl in a final volume of 25 μl) was subjected to PCR. The primers used and the PCR procedures have been described previously.11–15,16
RT-PCR was performed on RNA samples from all six malignant mesothelioma cell lines. PCR products were visualised by ethidium bromide staining of 1% agarose gels. Integrity of RNA and cDNA synthesis was monitored by amplification of β-actin mRNA (not shown).
Preparation of serum free conditioned medium
An identical number of SPC212 cells was seeded into 25 cm2 flasks and cultured until subconfluent. Serum supplemented medium was removed and the cell monolayer was extensively washed with PBS in order to remove remaining serum proteins. The cells were then incubated in serum free RPMI 1640 medium for 6 hours. Different growth factors were then added at the concentrations indicated and the cells were cultured for 36 hours. In some experiments the cells were preincubated with the tyrosine kinase inhibitor genistein (10 μg/ml) for 30 minutes and then treated with growth factors. Serum free conditioned medium from the growth factor treated and non-treated cells was collected, dialysed for 48 hours against distilled water, lyophilised, and used for zymography as described below.
Substrate zymography
Aliquots of the serum free conditioned medium (adjusted to the same number cells) treated or not treated with growth factor were solubilised in sample buffer containing 2% SDS without reducing agents. The samples were then applied without boiling on 6% polyacrylamide gels containing different substrates: 2 mg/ml gelatin and 0.5 mg/ml fibronectin, respectively. After electrophoresis the gels were washed in Hank’s balanced salt solution (HBSS) in the presence of 2.5% Triton X-100 for 2 × 20 minutes to remove SDS, then briefly rinsed in HBSS. The gels were then incubated in buffer (20 mM glycine, 10 mM CaCl2 and 1 μM ZnCl2, pH 8.3) for 48 hours at 37°C. After incubation the gels were stained in 0.1% Coomassie brilliant blue for 1 hour and destained. The substrate degrading enzymes were identified as clear bands in the blue background. The area and intensity of the bands was quantified by NIH image (1.60) software.
RESULTS
Expression of growth factor receptors in malignant mesothelioma cells by RT-PCR
To investigate potential molecular mechanisms responsible for regulation of MMPs by growth factors, we examined the mRNA expression of those growth factor receptors in malignant mesothelioma cells by RT-PCR.
Table 1 summarises the RT-PCR data on mRNA expression for the various growth factor receptors in six malignant mesothelioma cell lines. EGF receptor, c-met, IGF-I receptor, IGF-II receptor, c-kit and FGF receptor 4 was detected in all malignant mesothelioma cell lines; c-erbB4, PDGF receptor β and FGF receptor 3 was detected in 5/6 cell lines (except SPC212 and ZL5, respectively); mRNA for FGF receptor 1 was present in 4/6 cell lines; FGF receptor 2 was detected in only one cell line (STAV-FCS); and c-erbB2 and c-erbB3 mRNA could not be detected in any of the malignant mesothelioma cell lines.
Expression of growth factor receptors in malignant mesothelioma cells
Effects of growth factors on MMP-9 and MMP-2 activity in malignant mesothelioma cells
To investigate the effects of different growth factors on MMP-9 and MMP-2 activity in malignant mesothelioma cells, gelatin zymography was performed.
Aliquots of the growth factors treated or not treated with serum free conditioned medium from malignant mesothelioma cell SPC212 were subjected to SDS-PAGE gelatin zymography to examine the proteolytic activity. As shown in fig 1, the level of MMP-9 activity increased 1.9–8-fold on stimulation with EGF, TGF-α, AR, HB-EGF, BTC, IGF-I, HGF, aFGF and bFGF, respectively, in a concentration dependent manner compared with the unstimulated control. However, SCF, IGF-II and PDGF-BB had no effect or caused only a slight increase in MMP-9 activity (data not shown). In contrast, the level of MMP-2 activity was not affected by any of the growth factors investigated, indicating that MMP-2 production is not influenced by growth factor treatment. The absence of a change in MMP-2 activity levels indicates that the increase in MMP-9 can be attributed to growth factor stimulated production and not simply to a higher protein content in the samples. Human fibrosarcoma cell line HT-1080 was used as a positive control in all gelatin zymography studies.7

Effect of growth factors on the secretion of MMP-9 and MMP-2 in malignant mesothelioma cell line SPC212 by gelatin zymography. The gels show gelatinolytic activity in serum free conditioned medium in the presence or absence of different growth factors. Human fibrosarcoma cell line HT-1080 was used as a positive control for MMP-9 and MMP-2 in all gelatin zymography studies. Bar columns summarise the ratios of the area and intensity of MMP-9 and MMP-2 bands, as determined by image analysis, with 0 ng/ml MMP-9 band set at 1. The value is the mean (SD) of three separate experiments.
Effects of growth factors on MMP-3 activity in malignant mesothelioma cells
To investigate the effects of the same growth factors on MMP-3 activity in malignant mesothelioma cells, fibronectin zymography was performed.7
Aliquots of the growth factor stimulated serum free conditioned medium which gave maximum degradation of gelatin were subjected to fibronectin zymography. As shown in fig 2, the level of MMP-3 activity increased 1.7–5.3-fold when mesothelioma cells were stimulated with EGF, TGF-α, AR, HB-EGF, BTC, HGF, IGF-II, SCF and bFGF, respectively, compared with the unstimulated control. In contrast, IGF-I, PDGF-BB and aFGF had no effect or caused only a slight increase in MMP-3 activity in SPC212 cells.

Effect of growth factors on the secretion of MMP-3 in malignant mesothelioma cell line SPC212 by fibronectin zymography. The gel shows proteolytic activity in serum free conditioned medium in the presence or absence of different growth factors. The molecular weight (in kDa) is shown on the left. Bar column summarises the ratios of the area and intensity of MMP-3 bands, as determined by image analysis, with 0 ng/ml MMP-3 band set at 1. The value is the mean (SD) of three separate experiments.
Inhibitory effects of genistein on MMP activity in malignant mesothelioma cells
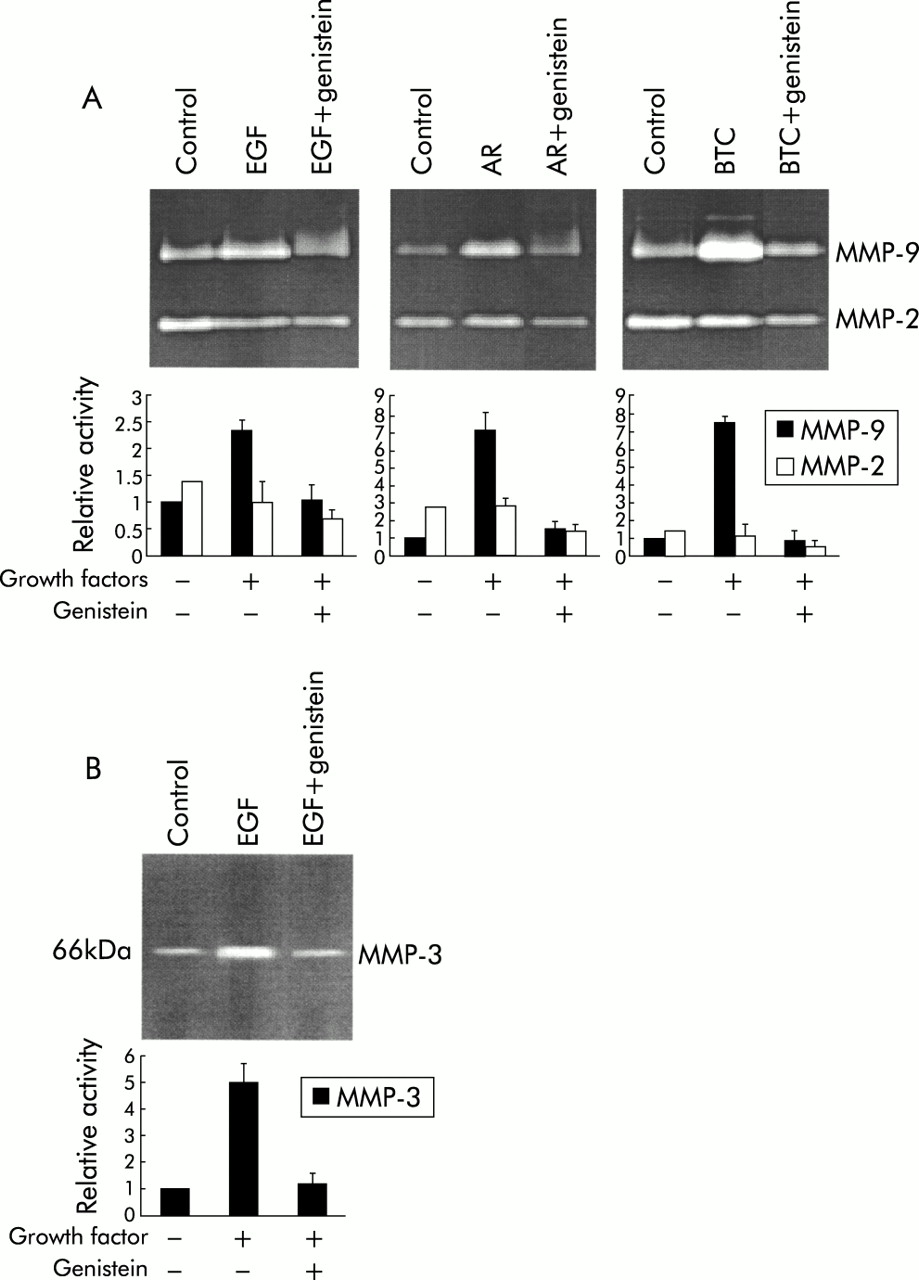
To investigate whether the tyrosine kinase intracellular pathway is involved in the increased production of MMPs by growth factors, SPC212 cells were preincubated with the protein tyrosine kinase inhibitor, genistein. As shown in fig 3, the increased production of MMP-9 and/or MMP-3 stimulated by the tyrosine kinase receptor ligands (EGF, AR and BTC; data for others not shown) was completely inhibited by genistein (10 μg/ml).
{kind=link}
{kind=link}
{kind=link}
Inhibitory effect of the tyrosine kinase inhibitor genistein on the production of (A) MMP-9 and (B) MMP-3 induced by different growth factors in malignant mesothelioma cell line SPC212. The gels show proteolytic activity in serum free conditioned medium in the presence or absence of genistein (10 μg/ml) and/or different growth factors (EGF 10 μg/ml, AR 50 μg/ml, and BTC 10 μg/ml). Bar columns summarise the ratios of the area and intensity of MMP-9 and MMP-2 bands or MMP-3 bands, as determined by image analysis, with 0 ng/ml MMP-9 or MMP-3 band set at 1. The value is the mean (SD) of three separate experiments.
DISCUSSION
Several studies have shown that the synthesis of MMPs is regulated by growth factors, cytokines, and cell contact with extracellular matrix in different tumour cells.6,8 For example, EGF and AR stimulate the expression of MMP-9 in breast cancer cells, fibronectin increases MMP-9 secretion in ovarian cancer cells, and EGF induces the expression of MMP-7 in prostate adenocarcinoma cells.18–20 However, the effect of various growth factors on secretion of MMPs in malignant mesothelioma cells has not been thoroughly investigated.
MMPs produced by tumour and stromal cells are believed to play a key role in tumour cell invasion and metastasis. We have shown that growth factors, including EGF, TGF-α, AR, HB-EGF, BTC, IGF-I, IGF-II, HGF, SCF, aFGF and bFGF, regulate the secretion of MMP-9 and/or MMP-3 in the human malignant mesothelioma cell line SPC212. In contrast, the secretion of MMP-2 was not affected by any of the growth factors investigated.
The regulation of MMPs is complex and occurs at both transcriptional and post transcriptional levels.8 Our findings are consistent with the differential transcriptional regulation of MMP-2, MMP-3 and MMP-9 by growth factors. This is due to the presence of different promoter elements on these MMP genes.21 The promoter of MMP-1, MMP-3, MMP-7 and MMP-9 contains an AP-1 element, a PEA3 element, and a TATA box consistent with induction by a variety of growth factors. The MMP-2 promoter is notably different from other MMP promoters, lacking the AP-1 element, the PEA3 element, and a classical TATA box. The growth factors may induce the expression and/or activation of c-Fos and c-Jun proto-oncogene products (which heterodimerise and bind AP-1) and Ets transcription factors (which bind PEA3) via the mitogen activated protein kinase (MAPK) pathways. Although regulation of the transcriptional level of MMPs is not directly studied here, our observation is in agreement with a previous study in which the expression of MMP-9, but not MMP-2, was found to be primarily regulated at the transcriptional level by growth factors.22 We speculate that similar mechanisms could be operational in the induction of MMP-9 and MMP-3 in malignant mesothelioma cells.
The growth factors investigated in our study increased the secretion of MMP-9 and MMP-3 in SPC212 cells to a varying degree. Beta-cellulin, AR, TGF-α and HB-EGF had the greatest stimulatory effect on MMP-9 secretion (increases of 8.0-fold, 6.7-fold, 4.9-fold, and 4.5-fold, respectively), whereas EGF had the greatest effect on MMP-3 secretion (5.3-fold increase). In contrast, SCF, IGF-II and PDGF-BB had only a slight or no stimulatory effect on MMP-9. IGF-I, PDGF-BB and aFGF had a similar effect on MMP-3 production. It should be noted that only PDGF-BB had a slight or no stimulatory effect on either MMP-9 or MMP-3. In addition, none of the examined growth factors stimulated production of MMP-2. Stable levels of MMP-2 in our experiments can also be viewed as an internal control. Increasing levels of MMP-9 and concomitant stable levels of MMP-2 confirm that the increase in MMP-9 must be a specific growth factor effect rather than the result of a larger amount of protein being loaded onto the gel.
The mechanism by which different growth factors regulate the expression of different MMP members is not fully understood. However, the signalling pathway initiated by growth factor receptor ligation which specifically interacts with the responsible growth factor is essential for signal transduction and biological function. Thus, whether or not those growth factor receptors are expressed in malignant mesothelioma cells may provide a potential mechanism for the binding of their ligands. To address this question we examined the mRNA expression of growth factor receptors in malignant mesothelioma cells by RT-PCR. Our results showed that all the malignant mesothelioma cell lines investigated expressed EGF receptor and five of the six cell lines (including SPC212) expressed c-erbB4, but none of the cell lines investigated expressed c-erbB2 and c-erbB3 which belong to the c-erbB receptor family. The c-erbB receptor family has at least eight cognate ligands including EGF itself, TGF-α and AR which only bind EGF receptor, BTC and HB-EGF which bind EGF receptor and erbB4, and several differentially spliced variants of neuregulin.23 Our finding that BTC and HB-EGF were the most effective in increasing MMP-9 secretion may be explained by their high affinity and stimulation of the tyrosine phosphorylation of either EGF receptor or erbB4.24
In agreement with our findings, previous studies have shown that most malignant mesothelioma cell lines express PDGF receptor-β which interacts with PDGF-BB.14 However, the mRNA expression of PDGF receptor-β could not be detected in malignant mesothelioma cell line SPC212. The lack of a stimulating response to PDGF-BB on both MMP-9 and MMP-3 induction may be explained by the absence of PDGF receptor-β expression on SPC212 cells.
We also found that most malignant mesothelioma cell lines expressed c-met, IGF-I receptor, IGF-II receptor, c-kit and four different FGF receptors, which is consistent with the induction of MMP-9 and/or MMP-3 by their ligands.
A number of studies have shown that several growth factors affect different aspects of cellular behaviour such as proliferation, motility, differentiation, and production of extracellular matrix proteins.11,18 Previous studies and our unpublished results have shown that malignant mesothelioma cells migrate to different growth factors.14,17 Furthermore, it has been shown that a number of growth factors stimulate the synthesis of the extracellular matrix molecule hyaluronan and proteoglycans in malignant mesothelioma cells.25 It has been indicated that increased production of hyaluronan by tumour cells or tumour associated fibroblasts is correlated with cell migration and metastasis.26 In our study we have shown that these growth factors also increase the secretion of MMP-9 and MMP-3 in malignant mesothelioma cells. These results strongly suggest that different growth factors may play an important functional role in the process of malignant mesothelioma cell invasion and metastasis.
Previous studies have shown that a tyrosine kinase pathway is involved in the signal transduction mechanisms of cell motility, the secretion of MMPs, and hyaluronan and proteoglycans production.11,27 The findings of our study are in agreement with these previous studies, since the increased production of MMPs promoted by different growth factors was inhibited by the tyrosine kinase inhibitor genistein.27 Selective chemical inhibitors for distinct signalling pathways can therefore be considered as targets for future therapeutic interventions.
In conclusion, this study has shown that exposure of the malignant mesothelioma cell SPC212 to different growth factors increases secretion of MMP-9 and MMP-3 but not MMP-2, and that BTC and EGF have the most potent effect on MMP-9 and MMP-3 production, respectively. Furthermore, there is evidence that the expression of different growth factor receptors in malignant mesothelioma cell lines corresponds to the stimulation of their ligands, and that the tyrosine kinase pathway is involved in the signal transduction mechanism. We have thus shown, for the first time, the broad extent of MMP regulation by a number of growth factors in malignant mesothelioma cells.
Acknowledgments
This work was supported by grants to JK from the Swedish Cancer Society and the Swedish Heart and Lung Foundation.