Article Text

Statistics from Altmetric.com
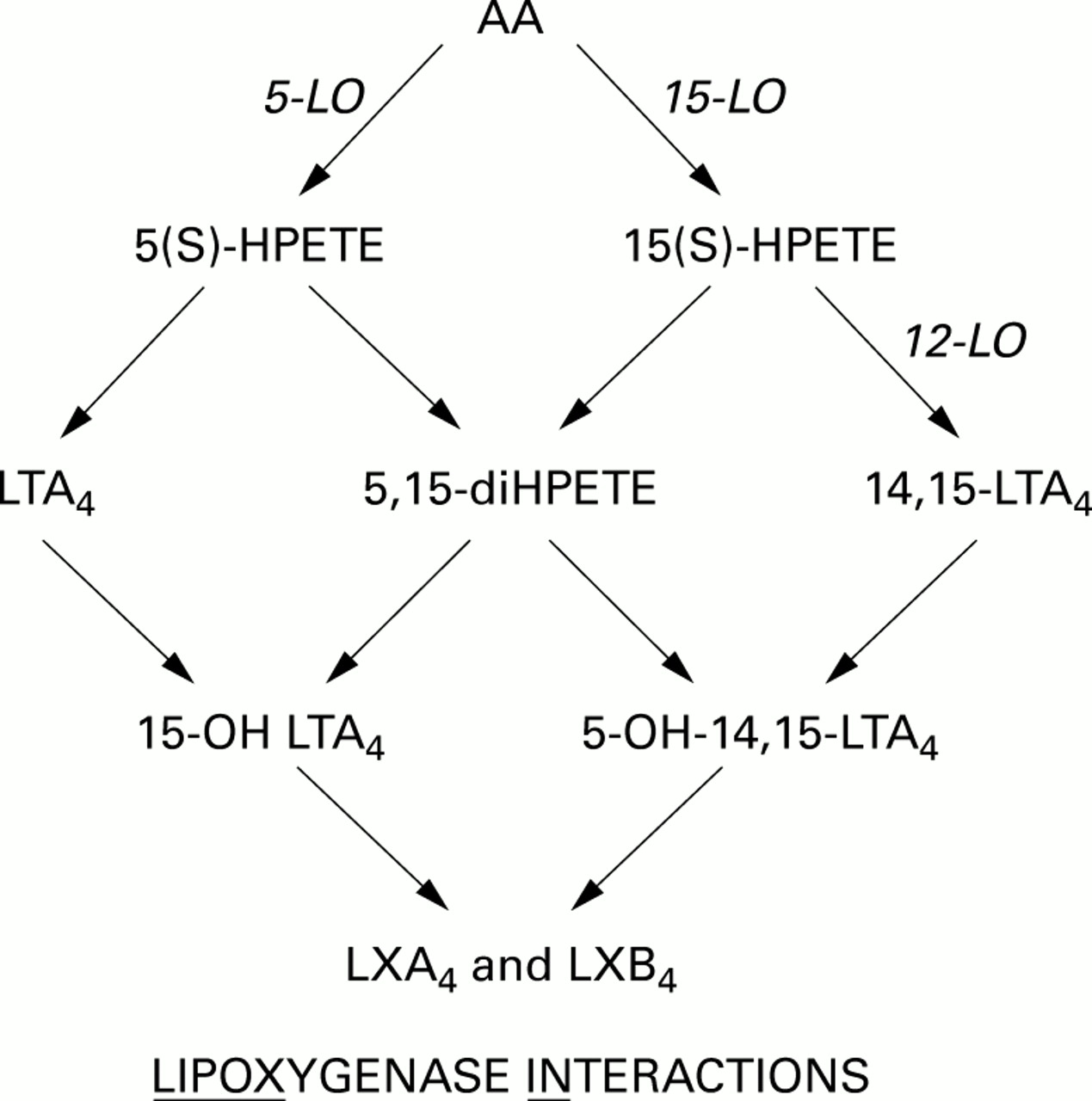
Leukotrienes and lipoxins, arachidonate derived mediators from the lipoxygenase (LO) pathways, are associated with bronchial asthma.1-3 They have been detected in vivo in bronchoalveolar lavage (BAL) fluid4-6 and are biosynthesised in vitro by human alveolar macrophages (AM) and polymorphonuclear cells (PMN) after non-specific stimulation.7 Lipoxins were first isolated by Serhanet al 8 ,9 who incubated granulocytes with 15(S)-hydroxyeicosatetraenoic acid (15(S)-HETE). Lipoxin synthesis was also achieved by the addition of other exogenous substrates to cell cultures.8 ,10 ,11 In contrast, lipoxin synthesis by co-cultures of two different cell types does not require the addition of any exogenous substrates.11-13 The biosynthesis of lipoxins was shown to be the result of a cellular cooperation mechanism between two different cell types. The enzymatic mechanism of transcellular metabolism involves the action of at least two different lipoxygenases (fig 1). Thus, lipoxins may be generated following receptor mediated activation of either co-incubated PMN and platelets,13 or ionophore stimulation of PMN alone after exposure to exogenous 15(S)-HETE.14
{kind=link}
Transcellular metabolism.
As shown previously,15 AM from asthmatic patients display high levels of 5-LO activity. 15(S)-HETE is considered to be specific to human epithelium airways and endothelial cells.16-18Human epithelial and endothelial cells are unable to produce lipoxins from endogenous 15(S)-HETE16 ,19 but they are surrounded by several cell types which show 5-LO activity. Moreover, lipoxins have been identified in the BAL fluid of patients with lung disease20 and in stimulated whole blood.21Because of these observations, and assuming that peripheral blood cells were activated before trafficking to the airways, we have investigated whether AM and peripheral blood cells display sufficient LO activity to transform 15(S)-HETE and have determined whether the generation of lipoxins and their precursors is related to the severity of asthma and can be modulated by treatment. Lipoxin A4(LXA4) has been shown to have a role in immune responses and to possess anti-inflammatory properties,22-24 and the presence of lipoxins in asthma has suggested that they impact on cell regulation. Glucocorticoids, commonly used in the treatment of inflammatory diseases, exert a number of controversial effects on the release of leukotrienes. Thus, lipoxins should be more specific biochemical markers of asthma and treatment efficacy than LTB4, and may be of interest for exploring glucocorticoid effects on cellular signalling and evaluation of long term treatment.
AM, PMN, and peripheral blood monocytes (PBM) from healthy controls, untreated asthmatics, and patients with steroid dependent asthma were studied for their ability to synthesise lipoxins and their precursors after stimulation with A23187 alone to investigate eicosanoid generation from endogenous arachidonate, and then in the presence of 15(S)-HETE to investigate transcellular metabolism.
Methods
PATIENTS
Asthma was diagnosed according to the criteria of the American Thoracic Society.25 The forced expiratory volume in one second (FEV1) ranged from 53% to 102% of the predicted values in patients with untreated asthma and from 45% to 117% of the predicted values in those treated with steroids. In the untreated asthmatic patients theophylline and β agonists were stopped one week before analysis. Non-steroidal anti-inflammatory drugs (NSAIDs) and corticosteroids had been discontinued for at least one month. The steroid dependent patients were included on the basis of their long term treatment (at least one year with oral prednisolone and inhaled beclomethasone) and their continued requirement for oral steroids.
PREPARATION OF CELLS
AM obtained by BAL centrifugation (400g, 10 min) from four control subjects and seven with untreated asthma were processed and cultured as previously described.26 PBM from nine control subjects, 18 with untreated asthma, and 29 steroid dependent asthmatics and PMN from nine controls and 38 untreated asthmatic subjects were isolated and purified from heparinised venous blood by centrifugation (400g, 20 min, 4°C) over a discontinuous Percoll gradient.27 ,28 Cell purity was greater than 95% and viability greater than 90%.
STIMULATION PROCEDURES AND METABOLITE ANALYSIS
After the addition of A23187 (5 × 10–6 M) which optimises eicosanoid production, with or without 15(S)-HETE (3 × 10–6 M), incubations were carried out for five minutes for PMN (107 cells/ml) and for 30 minutes for AM (106 cells/ml) and PBM (5 × 106cells/ml). The samples were analysed using reverse phase high performance liquid chromatography (RP-HPLC).27-29
ANALYSIS OF DATA
The results are presented as mean (SE) ng product/5 × 106 cells. Statistical differences were determined using the Mann-Whitney U test for unpaired samples. A p value of <0.05 was considered significant.
Results
EICOSANOID RELEASE FROM ENDOGENOUS SOURCES
Cell capacity to release eicosanoids from endogenous arachidonic acid was evaluated using ionophore A23187 alone. AM, PMN, and PBM released LTB4.27-29 No detectable amounts of diHETEs and lipoxins were observed under our analysis conditions in AM or PBM. Nevertheless, besides the normally generated leukotrienes, PMN from untreated asthmatic subjects were able to generate considerable amounts of 5(S),15(S)-diHETE as well as lipoxins (135 (14) and 6 (1), respectively). These metabolites were not found in PMN from control subjects.
EICOSANOID RELEASE FROM TRANSCELLULAR METABOLISM
Transcellular metabolism was evaluated by studying 15(S)-HETE metabolism in A23187 stimulated cells. LTB4, lipoxins, and their precursor 5(S),15(S)-diHETE were detected in all three cell types, whereas 14(R),15(S)-diHETE was released by PBM only (table1).27-29 5(S),15(S)-diHETE, which was the major product, was at a similar level in AM from patients with untreated asthma and healthy controls. However, it was nearly twofold higher in PMN from patients with untreated asthma than in those from healthy controls,27 whereas no differences were seen in PBM.28 Total lipoxin levels were significantly higher in the cells from untreated asthmatic subjects than from healthy control subjects for the three cell types. 14(R),15(S)-diHETE levels, generated by PBM only, were almost identical for the two groups of subjects.
Eicosanoid generation from transcellular metabolism
EFFECTS OF ORAL CORTICOSTEROIDS ON TRANSCELLULAR METABOLISM
PMN from steroid dependent asthmatics released twofold lower levels of 5(S),15(S)-diHETE than those from untreated asthmatics and a similar level to those from controls (table 1). Cells from patients with steroid dependent asthma also released lower lipoxin levels than those from untreated asthmatics and controls. In contrast, PBM from patients with steroid dependent asthma released higher levels of 5(S),15(S)-diHETE than those from untreated asthmatic patients and controls, whereas lipoxin levels were nearly the same. Simultaneously, PBM from patients with steroid dependent asthma released significantly greater amounts of 14(R),15(S)-diHETE than those from patients with untreated asthma or healthy controls.
Discussion
Human cells were studied ex vivo for their ability to synthesise eicosanoids as an indication of cellular activation in vivo. They were able to synthesise lipoxins and their precursors 5(S),15(S)-diHETE or 14(R),15(S)-diHETE. In our studies lipoxins were released from endogenous sources in PMN from untreated asthmatic patients only. Nevertheless, Levy et al described lipoxin synthesis in the sub-nanogram range in cytokine primed human AM suspensions using an electrochemical detection technique.30 Endogenous generation of 5(S),15(S)-diHETE and lipoxins in vitro by human blood PMN, without in vitro cytokine priming, has been reported for the first time in rheumatoid disease by Thomas et al 31 and later in asthma,27 although other authors have since claimed that only porcine leucocytes were able to produce lipoxins from endogenous arachidonate.32 It was the first time that lipoxin biosynthesis was described in a cell type with the involvement of 5-LO only. These results can be interpreted by in vivo priming induced in bronchial diseases by cytokines or tumour necrosis factor (TNF)-α released by surrounding cells.13
In contrast, lipoxins were produced in all cell types when incubated with exogenous mediator 15(S)-HETE.27-29 ,31 AM, PMN, and PBM from untreated asthmatic subjects produced greater quantities of lipoxins and their precursors than those from healthy controls by way of transcellular metabolism. These results suggest that the more active transcellular metabolism of phagocytes from patients with untreated asthma may be associated with previous exposure to specific allergens responsible for the inflammatory process and induced by subsequent lung disease. This may indicate that lipoxin can be produced at inflammation sites from a massive amount of 15(S)-HETE generated by airway epithelium and/or human bronchi.16 ,19 This could explain the presence of LXA4 in BAL fluid from patients with lung diseases involving a cellular cooperation mechanism, thus leading to mediators playing an important part as immunoregulatory molecules in asthma.20
It is commonly accepted that 5-LO activity is reduced in the presence of glucocorticoids. We observed that lipoxin and 5(S),15(S)-diHETE levels released by PMN from steroid dependent asthmatics by way of transcellular metabolism were lower than those from patients with untreated asthma and healthy controls, and were completely inhibited in the endogenous pathway. In contrast, the amounts of diHETEs biosynthesised from exogenous 15(S)-HETE were increased in PBM from patients with steroid dependent asthma compared with those from untreated asthmatics and control subjects, with significant differences for 14(R),15(S)-diHETE. Although lipoxin levels are nearly the same in PBM from both untreated and steroid dependent asthmatic patients, our results have shown that they are not synthesised in control subjects. This result is unexpected since glucocorticoids are thought to reduce lipid mediator production by their anti-inflammatory effects. The role of glucocorticoids in the regulation of 5-LO activity is uncertain. Studies performed in vitro did not show a significant effect on LTB4 release by human blood neutrophils33 and monocytes,34 whereas inhibition of specific eicosanoid synthesis and induction of lipocortin-1 by glucocorticoids were reported for macrophages.35 In contrast, a recent study reported increased expression of 5-LO in blood monocytes from healthy volunteers cultured in the presence of dexamethasone,36and glucocorticoid mediated upregulation of the neuronal 5-LO pathway has been described in rats treated with corticosterone or dexamethasone.37
Our study shows an upregulation of LO activity in PBM following long term treatment with corticosteroids, and lipoxins have been found to regulate leucocyte trafficking in the vascular circulation by inhibition of several inflammatory responses.24 These results suggest that the anti-inflammatory properties of glucocorticoids and the clinical improvement of patients might result in part from the use by target cells of an endogenous anti-inflammatory mediator released by the surrounding cells. Lipoxins and their precursors could be of interest for explaining the effects of glucocorticoids on cellular signalling and for evaluating long term treatments.