Article Text

Abstract
BACKGROUND Broncho-Vaxom (OM-85 BV) is known to support respiratory tract resistance to bacterial infections. In vivo and in vitro studies in animals and humans have shown that the action of the drug is based on the modulation of the host immune response, and it has been found to upregulate interferon γ (IFN-γ) and interleukin (IL)-2, IL-6, and IL-8. These immunomodulatory effects of the compound may explain its stimulation on T helper cells and natural killer cells. Following earlier findings that OM-85 BV induces the synthesis of IL-6, a study was undertaken to investigate its possible effect on other gp130 binding cytokines including IL-11, IL-12, leukaemia inhibitory factor (LIF), oncostatin M (OSM), and ciliary neutrophil factor (CNTF). Its modulation of the corresponding receptors of the above mentioned cytokines and of the signal transducer gp130 in human pulmonary fibroblasts and peripheral blood lymphocytes was also studied.
METHODS Transcription of cytokines was assessed by Northern blot analysis. Secretion of cytokines was analysed using commercially available enzyme linked immunosorbent assay kits. Cytokine receptors and gp130 proteins were determined by Western blot analysis.
RESULTS OM-85 BV increased the expression of IL-11 in human lung fibroblasts, but not in lymphocytes, in a dose and time dependent manner by maximal fivefold within 20 hours. The compound inhibited serum induced IL-12 expression in peripheral blood lymphocytes but did not induce OSM, LIF, or CNTF at any concentration. In lung fibroblasts the expression of the IL-6 receptor was enhanced fourfold at a concentration of 10 μg/ml OM-85 BV while that of the IL-11 receptor was not altered. In peripheral blood lymphocytes LIF receptor α expression was downregulated in the presence of 10 μg/ml OM-85 BV. At a concentration of 10 μg/ml OM-85 BV enhanced gp130 gene transcription fivefold and increased gp130 protein accumulation in cell membranes by 2.5 times.
CONCLUSION In vitro OM-85 BV exerts immunomodulatory action via modulation of the signal transducer gp130 and gp130 binding cytokines. The increase of IL-6 and IL-11 may explain enhanced T and B cell activity, immunoglobulin synthesis, and IgM to IgG switch. Suppression of IL-12 and LIF receptor-α further contributes to organ protection. With regard to gp130 mediated signalling of the investigated cytokines, OM-85 BV modifies the host immune response towards an increased sensitisation of cells to gp130 binding proteins.
- Broncho-Vaxom (OM-85 BV)
- gp130
- signal transduction
Statistics from Altmetric.com
Broncho-Vaxom (OM-85 BV; OM Laboratories, Geneva, Switzerland) is a lyophilised extract of eight species of bacteria which frequently cause inflammation of the respiratory tract. The compound has been shown to modulate host immune responses leading to increased resistance against bacterial infections.1-5 Enhanced antibody synthesis together with better resistance to bacterial infection was documented when OM-85 BV was administered orally.1-6
The protective effect of OM-85 BV against acute respiratory exacerbations has recently been shown in a large double blind, placebo controlled, randomised clinical study in 381 patients with chronic obstructive pulmonary disease (COPD), 190 of whom were give placebo and 191 OM-85 BV. The risk of an acute exacerbation was similar in both groups. However, the number of days spent in hospital with respiratory problems was reduced by 55% in the group treated with OM-85 BV compared with the placebo treated patients. In addition, dyspnoea improved in the patients treated with OM-85 BV but deteriorated slightly in patients receiving placebo. These results suggest that OM-85 BV may be beneficial for patients with COPD.2 OM-85 BV has been shown to increase polymorphonuclear leucocytes and to impair the activity of alveolar macrophages in local airway immunity. An analysis of pre- and post-therapeutic bronchoalveolar lavage (BAL) fluid of 28 adult patients with non-obstructive chronic bronchitis revealed that OM-85 BV increased the CD4+/CD8+ lymphocyte ratio and IFN-γ levels. Furthermore, the alveolar macrophage activity was normalised and the BAL IgA was moderately increased. In earlier studies it has been reported that OM-85 BV enhances natural killer cell activity and increases the spontaneous and phytohaemagglutinin induced production of tumour necrosis factor (TNF)-α and IL-2. It also increases phytohaemagglutinin stimulated production of IFN-γ.5 The efficacy of OM-85 BV in recurrent respiratory and ear, nose and throat infections in 61 children has been compared with placebo treatment in 55 children.6 This double blind, randomised study followed a 90 day treatment period and a 90 day follow up period without drugs; 39.5% of all children taking OM-85 BV remained free from infection compared with 16.5% in the placebo group. In the OM-85 BV group the use of antibiotics was reduced by 44% compared with a reduction of 23.5% in the placebo group.6 Similar results of reduced use of antibiotics were reported by Debbas and Derenne7 in 265 elderly patients suffering from various infectious diseases of the upper respiratory tract.
The mechanism by which OM-85 BV activates the host immune response is not completely understood. Animal studies suggest that antigenic proteins contained in OM-85 BV may be passed unchanged from the gut to the lymphatic system.8 Using an animal model the compound has been shown to activate macrophage and polyclonal B lymphocytes. It exerted an immunogenic effect by inducing serum IgG binding to the bacterial strains used for the preparation of OM-85 BV.9Serum probes obtained from OM-85 BV treated animals recognised specific bacterial cell wall components including porin, lipoprotein/lipopeptide, and murein. In addition, OM-85 BV exhibited adjuvant properties when applied with antigens such as TNP-BSA or an influenza vaccine.9
An investigation of the underlying molecular biological mechanisms of the above described in vivo and in vitro studies showed that OM-85 BV can induce the expression of IL-6 in cultures of human pulmonary fibroblasts.10 Recent studies support the idea that IL-6 acts as a protective rather than a pro-inflammatory factor,11-14 and diminishes tissue inflammation in hypersensitised pneumonitis,12 oxygen toxicity,13 and endotoxin induced lung injury.14
IL-6 exerts its biological activity by binding to a specific 80 kDa IL-6 receptor which contains no intracellular signalling domain.15 ,16 To induce IL-6 dependent cell activation the IL-6/IL-6 receptor complex has to bind to another membrane bound signalling protein, gp130.15 ,16 This functional complex can be formed by either form of the IL-6 receptor—the membrane bound form or the soluble form. Some cell types are not able to synthesise the IL-6 receptor but react to IL-6 in the presence of the soluble IL-6 receptor.15 ,16
Interestingly, gp130 serves as signal transducer for several other cytokines including leukaemia inhibitory factor (LIF),17 ,18 oncostatin M (OSM),17ciliary neutrophil factor (CNTF),17IL-11,16 ,18 and probably IL-12.16 ,19 The signal transduction complexes formed by gp130 with either LIF, OSM, CNTF, and IL-11 appear to involve a cytokine/cytokine receptor complex similar to that described for IL-6/IL-6 receptor.16 ,17Since all gp130 binding cytokines affect host immune responses, we have investigated the possible effect of OM-85 BV on the expression of the above mentioned factors, their specific receptors, and on gp130 in cultivated human pulmonary fibroblasts and peripheral blood lymphocytes.
Methods
DRUG
OM-85 BV is a lyophilised bacterial lysate consisting ofHaemophilus influenzae,Diploccocus pneumoniae,Klebsiella pneumoniae,Klebsiella ozeanae,Staphylococcus aureus,Streptococcus pyogenes,Streptococcus viridans, andNeisseria catarrhalis.
CULTURE OF HUMAN PULMONARY FIBROBLASTS
Cultures of primary human lung fibroblasts were established from sterile lung biopsy specimens obtained from six patients undergoing pneumonectomy for other reasons. Cells were grown from small pieces of tissue (5 × 5 mm) in a medium composed of RPMI 1640 supplemented with 8 mM stabilised l-glutamine, 10% fetal calf serum, and 20 mM HEPES.20 The same medium was used for subsequent cultures of cells. No antibiotics or antimycotics were added throughout the entire cell culture. Cells were characterised by their morphological appearance on light microscopy and by immunofluorescence analysis.20 All experiments were carried out before the seventh passage.
STIMULATION OF FIBROBLASTS
Subconfluent layers of fibroblasts (80% density) were starved by serum depletion (RPMI 1640 + 0.5% fetal calf serum, 8 mM stabilisedl-glutamine, 20 mM HEPES) for 48 hours and the medium was exchanged every 12 hours during the period of starvation. Cells were stimulated with various concentrations of sterile filtered OM-85 BV (0.1–10 μg/ml) for various time periods (0, 6, 12, 24, 36, and 48 hours) before they were either lysed for analysis of RNA, scraped for protein analysis, fixed for immunostaining, or before culture medium was collected for quantitative determination of the various cytokines by enzyme linked immunosorbent assays (R&D Systems).
ISOLATION OF PERIPHERAL BLOOD LYMPHOCYTES
Ten ml peripheral blood was obtained from four healthy volunteers and diluted 1:5 with ice cold phosphate buffered saline (PBS; Seromed/Fakola). The diluted blood was layered onto 20 ml Ficoll (Sigma) and lymphocytes were isolated by centrifugation at 600g for 30 minutes at 4°C. Lymphocytes were carefully removed from the Ficoll layer and washed twice with 10 ml PBS (600g for 30 minutes at 4°C). The lymphocytes were adjusted to a cell density of 104/ml cell culture medium (RPMI 1640 + 10% fetal calf serum, 8 mM stabilised l-glutamine, 20 mM HEPES). Stimulation was performed as described for fibroblasts.
NORTHERN BLOT ANALYSIS
Total RNA was extracted by Trizol (Gibco/BRL). Ten μg of total RNA was applied to a denaturing 1% agarose gel containing 5% formaldehyde. After size fractionation, RNA was blotted onto a nylon membrane (Amersham) by capillary transfer overnight in 10 × SSC buffer (1 × SSC: 0.156 M NaCl, 0.014 M Na citrate).10RNA was then immobilised and bound to the nylon membrane by ultraviolet irradiation. Membranes were prehybridised using QuickHyb buffer (Stratagene) for 30 minutes at 60°C. Probing for the mRNAs of the various cytokines was achieved using commercially available [α32P]dATP 5′-end labelled oligonucleotides (all cDNA probes from R&D Systems). Hybridisation was performed after addition of one of the labelled oligonucleotides at 55°C for two hours using the same buffer. After twice washing in 1 × SSC at room temperature a third wash was performed at 50°C for 15 minutes. The membranes were finally washed once in 0.5 SSC at 50°C for 10 minutes and then exposed to a BioMax film (Kodak) overnight. Northern blot analysis was performed using a computer assisted image analysis program. A random prime labelled HLA-β cDNA probe was used as a control in all experiments.10 ,21 ,22
ENZYME LINKED IMMUNOSORBENT ASSAYS
Following stimulation, equal amounts (1 ml) of cell culture medium were collected at the time points indicated above. Secreted proteins of LIF, OSM, IL-11, and IL-12 were determined in the culture medium using the respective commercially available enzyme linked immunosorbent assays (all from R&D Systems) following the methods advised by the manufacturer.
WESTERN BLOT ANALYSIS
Proteins of serum starved and OM-85 BV stimulated cells were harvested by scrapping the cell layer after being washed twice with ice cold PBS. Cells were resuspended in a denaturing Laemmli SDS lysis buffer (62.5 mM Tris HCl, pH 6.8; 2% SDS, 0.5% mercapto-ethanol, 10% glycerol, 0.01% bromophenol blue) and the protein concentration was determined by Lowry assay.21 Equal amounts of protein were applied onto a denaturing 4–15% gradient SDS-polyacrylamide gel (SDS-PAGE) and the proteins were size fractionated (45 mA) for two hours. The proteins were then transferred onto a PVDF membrane (Amersham) and non-specific binding of antibodies was prevented by incubation of the membranes for one hour in PBS supplemented with 5% skimmed milk (blocking buffer). Following membranes were incubated in blocking buffer in the presence of the respective first monoclonal antibody (gp80, MAB227; IL-11 receptor, AF490; LIF receptor-α, AF-249-NA; all from R&D, dilution 1:1000) for two hours at room temperature. Unbound antibodies were removed by three washes with blocking buffer before membranes were incubated overnight with a second peroxidase labelled anti-mouse antibody in blocking buffer (R&D, dilution 1:1000). Non-bound antibody was removed by two washings with blocking buffer and the protein bands were visualised using a chemiluminescence kit (Amersham) in combination with BioMax film (Kodak).21
STATISTICAL ANALYSIS
The null hypothesis for all experiments was that there is no difference between untreated cells and cells exposed to OM-85 BV. This was analysed by ANOVA for all experimental data.
Results
At therapeutic concentrations OM-85 BV induced the expression of IL-11 in six primary human pulmonary fibroblasts in a time and dose dependent manner. All cell lines responded in a similar pattern to the presence of the compound. A representative trace of OM-85 BV induced IL-11 mRNA synthesis is shown in fig 1A. The increase in IL-11 mRNA synthesis started after four hours of incubation and reached a maximal level at six hours, persisting for the entire period of incubation (fig1A). The dose dependent induction of IL-11 mRNA by OM-85 BV was analysed 12 hours after addition of the compound. The induction of IL-11 gene transcription was observed at concentrations of >1 μg/ml and achieved a maximal effect at 10 μg/ml (fig 1B). Both time and dose dependent effects of OM-85 BV on transcription of IL-11 varied for ±2 hours in different cell lines. Induction of IL-11 mRNA synthesis was reduced when cells were pretreated with actinomycin D (5 μg/ml), which suggests that the increase in IL-11 mRNA signals was due to de novo synthesis rather than to accumulation of mRNA by delayed degradation (data not shown). In contrast, OM-85 BV did not induce IL-11 mRNA synthesis in peripheral blood lymphocytes (data not shown). Experiments were performed in all six fibroblast cell lines at each time point and dose in triplicate.

(A) Representative trace of OM-85 BV (Broncho-Vaxom) showing induced de novo synthesis of IL-11 mRNA in human lung fibroblasts. HLA-β was used as constitutive house keeping gene. (B) Dose dependent induction of IL-11 mRNA synthesis in human lung fibroblasts. (C) Graph showing OM-85 BV induced secretion of IL-11 by six human fibroblast cell lines. Each data point represents the mean (SE) of six independent experiments each performed in triplicate and bars indicate standard error.
The enhancing effect of OM-85 BV on IL-11 mRNA was confirmed by an increase in secreted IL-11 in the cell culture medium obtained from the six fibroblast cell lines studied. Three independent medium samples were collected from individual cell culture wells of each of the six investigated cell lines at each time point. Increasing amounts of secreted IL-11 were observed 12 hours after the start of incubation (fig 1C). Statistical analysis using ANOVA revealed a significant increase in secreted IL-11 (p<0.01 at times longer than 12 hours). The changes in secreted IL-11 protein induced by 10 μg/ml OM-85 BV were confirmed by immunosorbent assay (fig 1C). The effect of the compound was clearly dose dependent (data not shown). Again, lymphocytes did not synthesis IL-11 in the presence of OM-85 BV.
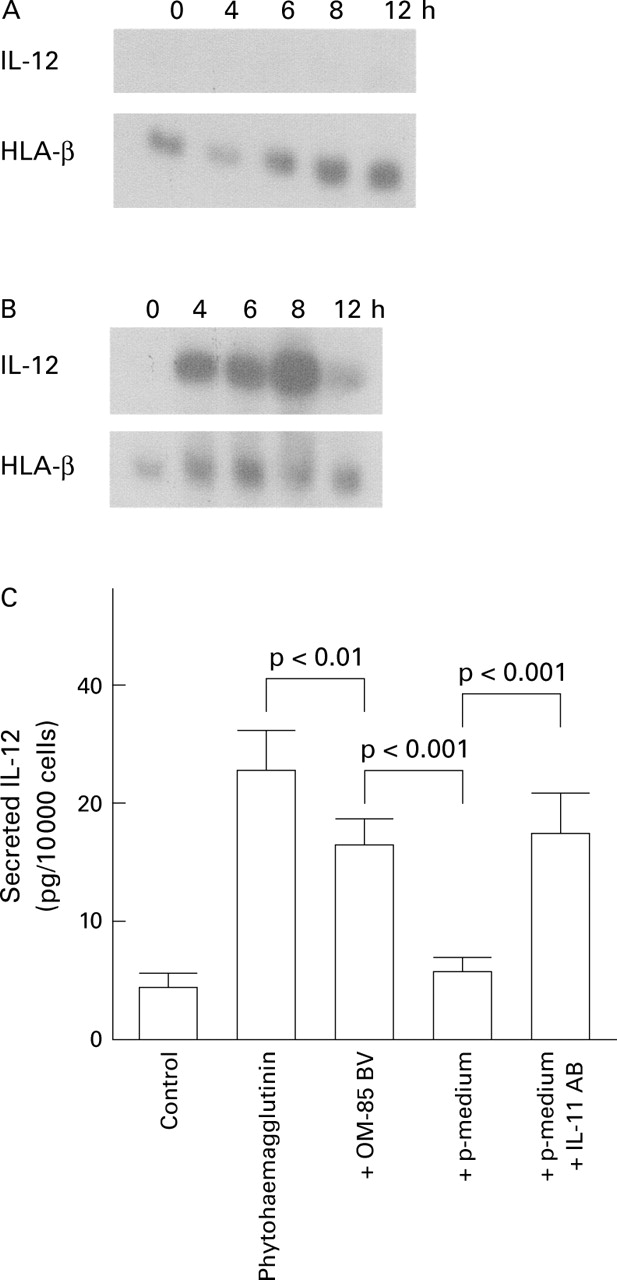
None of the six human pulmonary fibroblast cell lines synthesised IL-12 mRNA under any of the conditions investigated (fig 2A). In contrast, peripheral blood lymphocytes obtained from four healthy volunteers could be stimulated to synthesise IL-12 mRNA with either phytohaemagglutinin or 10% fetal calf serum (fig 2B). OM-85 BV alone did not induce IL-12 synthesis (data not shown). Phytohaemagglutinin induced a time dependent de novo synthesis of IL-12 mRNA as shown in fig 2B. Simultaneous application of OM-85 BV inhibited IL-12 synthesis by phytohaemagglutinin in peripheral blood lymphocytes in a dose dependent manner (data not shown).

(A) Kinetics of IL-12 mRNA synthesis induced by OM-85 BV (Broncho-Vaxom) in six human lung fibroblast lines. HLA-β was used as constitutive house keeping gene. (B) IL-12 mRNA synthesis by phytohaemagglutinin in peripheral blood lymphocytes. HLA-β was used as constitutive house keeping gene. A and B originate from the same gel and have been split for clarity. (C) Inhibitory effect of 10 μg/ml OM-85 BV on phytohaemagglutinin induced secretion of IL-12; p-medium = preconditioned medium obtained from fibroblasts incubated for 24 hours in the presence of 10 μg/ml OM-85 BV. Neutralising IL-11 antibodies (IL-11 AB) were used at a concentration of 50 μg/ml. The inhibitory effect of 10 μg/ml OM-85 BV on phytohaemagglutinin induced IL-12 synthesis was significant (p<0.01) and compared with preconditioned fibroblast medium (p<0.001). The anti-inhibitory effect of anti-IL-11 antibodies on p-medium in the presence of phytohaemagglutinin was significant (p<0.001). Each data point represents the mean (SE) of six independent experiments each performed in triplicate.
Incubation of blood lymphocytes with preconditioned culture medium obtained from fibroblasts pre-incubated for six hours with 10 μg/ml OM-85 BV resulted in inhibition of IL-12 synthesis. As shown in fig 2C, the phytohaemagglutinin induced expression of IL-12 in lymphocytes was downregulated to basal levels in the presence of fibroblast OM-85 BV preconditioned medium (p<0.001).
Based on a report that IL-11 downregulates IL-12 expression,22 we investigated the possible role of OM-85 BV induced IL-11 in fibroblast medium and its role on IL-12 expression. The effect of medium preconditioned with OM-85 BV on IL-12 inhibition could be counteracted by neutralising IL-11 antibodies dose dependently. The inhibitory effect of OM-85 BV preconditioned medium neutralised by 70% resembled the inhibitory effect observed with OM-85 BV alone (fig 2C). Experiments with preconditioned medium obtained from each of the six fibroblast cell lines were performed with each of the four lymphocyte samples from different donors.
OM-85 BV did not induce OM, LIF or CNTF in either human lung fibroblast cultures or in peripheral blood lymphocytes.
Since all the cytokines studied need complex formation with their specific receptors to bind to their shared signalling transducer gp130, we investigated whether OM-85 BV affects the expression of these receptors on both cell types. Western blot analysis revealed a rapid increase in IL-6 receptors in fibroblast lysates after stimulation with OM-85 BV (fig 3A). Both the membrane bound and soluble isoforms of the IL-6 receptor could be detected by Western blotting. The increase in both IL-6 receptors occurred synchronously; however, while expression of the membrane bound form of the IL-6 receptor appeared to be constant after six hours, that of the smaller soluble IL-6 receptor was increased over the whole observed time period (fig3A).

(A) Representative Western blot analysis of IL-6 receptor expression in human lung fibroblasts. (B) Representative Western blot analysis of the IL-11 receptor on peripheral blood lymphocytes. Similar results were obtained in human lung fibroblasts. (C) Downregulation of LIF receptor α on peripheral blood lymphocytes by OM-85 BV. (D) Densitometry analysis of the expression of IL-6 receptor, IL-11 receptor, or LIF receptor α proteins. Each data point represents the mean (SE) of six independent experiments each performed in triplicate.
Transcription of the IL-11 receptor gene was observed in unstimulated cells in both lung fibroblasts (n = 6) and peripheral blood lymphocytes (n = 4) and was only slightly increased by OM-85 BV at various concentrations ranging from 0 to 10 μg/ml (data not shown). Protein expression of the IL-11 receptor was only detectable at a low level and the OM-85 BV induced increase was undetectable before eight hours incubation (fig 3B). As shown in figs 3B and D, the OM-85 BV induced IL-11 receptor protein expression was increased during the entire time period observed. However, the effect of the drug was only observed at the highest concentration (10 μg/ml) in both cell types, assessed with all six fibroblast cell lines and in all four lymphocyte samples.
The LIF receptor α was expressed at a low level in peripheral blood lymphocytes (n = 4) but in none of the six fibroblast cell lines (fig3C). Interestingly, this receptor was downregulated by the presence of OM-85 BV. As shown in figs 3C and D, the effect was time dependent and the LIF receptor α was not detectable after 12 hours of incubation. As was the case with expression of the IL-11 receptor α protein, the inhibitory effect of OM-85 BV on LIF receptor was only observed at the highest concentration used. Western blot analysis of receptors for OSM, IL-12, and CNTF were negative.
All the cytokine receptors studied need to interact with their unique signal transducer gp130 to exert a cellular response. We therefore investigated the effect of OM-85 BV on the expression of the gp130 gene in human primary fibroblast cell lines (n = 6). As shown in fig 4A, we observed a low constitutive signal for the gp130 mRNA in unstimulated cells. OM-85 BV induced a constant increase in gp130 mRNA synthesis which remained at a high level of expression for the entire observation period (fig 4A).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Northern blot analysis of the effect of OM-85 BV on de novo synthesis of gp130 mRNA in human lung fibroblasts. HLA-β was used as a control house keeping gene. Similar results were obtained in peripheral blood lymphocytes. (B) Western blot analysis of the stimulatory effect of OM-85 BV on gp130 protein expression in human lung fibroblasts. (C) Densitometry analysis of gp130 mRNA and gp130 protein expression in human lung fibroblasts. Each data point represents the mean (SE) of six independent experiments each performed in triplicate.
The increase in gp130 mRNA induced by OM-85 BV was sustained by protein analysis by Western blotting using total cellular protein preparations. As depicted in a representative Western blot (fig 4B), the expression of the gp130 protein is similar to the data obtained by mRNA analysis, revealing a relatively high spontaneous expression of the gp130 protein in unstimulated cells which was increased by OM-85 BV (fig 4B). Analysis of Northern and Western blots by ANOVA revealed a significant increase in gp130 mRNA but a less prominent increase in gp130 protein synthesis in all six fibroblast cell lines (fig 4C). The effect occurred at a concentration of 5 μg/ml and was not detectable at lower concentrations (data not shown).
Discussion
We hypothesised that OM-85 BV exerts some of its immune modulatory effects via gp130 binding cytokines including IL-6, IL-11, IL-12, OM, ILF, and CNTF and may also affect gp130 itself. This hypothesis could be proved in part; while the compound activates two gp130 binding interleukins (IL-11 and IL-610), it downregulates IL-12. None of the other known gp130 activators was induced by OM-85 BV in human pulmonary fibroblasts. In addition, OM-85 BV upregulates the corresponding interleukin receptors—namely, the IL-6 receptor and the IL-11 receptor—and, most importantly, the compound increased the expression of the signal transducer gp130 in human peripheral blood lymphocytes and pulmonary fibroblasts. The leucocyte inhibitory factor (LIF) receptor was downregulated.
IL-6 has been suggested to be an organ protective factor rather than a pro-inflammatory factor.12-14 It has been shown to stimulate T cell growth, T cell dependent B cell maturation, megakaryopoiesis, and various stages of myeloid differentiation. On the other hand, IL-6 is known to inhibit adipogenesis.16 ,23 ,24 The action of IL-6 may therefore be regarded as organ protective and supportive for the host's immune response towards bacterial and viral infections.11 ,16 ,23-25 The controversial findings concerning the biological action of IL-6 on the immune system may depend on organ or disease specific physiological/pathophysiological conditions and on the priming of the respective tissues or cells studied.11 ,12 ,17 ,23-25
With regard to the protective immunomodulatory effects of IL-11, it is noteworthy that these are almost identical to those of IL-6.11 ,16 ,23-25 IL-11 has been shown to share many activities with IL-6. However, the activity of IL-11 on different stages of erythropoiesis in vitro clearly sets it apart from IL-6. IL-11 has little haematopoietic colony stimulatory capacity although it sustains terminal differentiation of the late erythroid progenitors.23 ,24 In combination with IL-3, IL-11 has profound stimulatory effects on early multipotent haematopoietic progenitors, on multi-lineage colony forming cells, as well as on erythroid progenitors.23-25 Furthermore, IL-11 increases the number of circulating neutrophils and platelets and increases megakaryopoiesis in normal mice and primates.23-26 In addition, IL-11 prevented lethal graft-versus-host disease in bone marrow transplant recipients.26 It also reduced serum levels of circulating TNF-α and suppressed TNF-α secretion by lipopolysaccharide (LPS) stimulated macrophages in vitro.26 Thus, IL-11 decreased morbidity and mortality.26 IL-11 minimises tissue injury by its ability to protect clonogenic stem cells, to regulate epithelial cell proliferation, inhibit apoptosis, and inhibit macrophage cytokine production.24 ,25 Thus, IL-11 appears to play an important role in haematopoiesis, bone metabolism, and tissue remodelling and may be an important protector of mucosal surfaces.24 However, as with IL-6, the immune modulatory effects of IL-11 depend on the stage of cell differentiation and microenvironmental conditions of the cells affected by IL-11.23-26 The interaction of interleukins, especially that of IL-6 and IL-11, has been found in human B cells which shows that IL-11 promotes differentiation of human B cells only in the presence of IL-6 activated accessory T cells and monocytes.25 With regard to our observations on the cytokine regulatory effect of OM-85 BV on human pulmonary fibroblasts and peripheral blood lymphocytes, these findings can explain some of the most important immunomodulatory action of the compound observed in clinical trials.
Together these observations define IL-6 as a factor that preferably triggers T and B cell proliferation while IL-11 seems to activate a gp130 mediated signalling cascade which induces B cell differentiation resulting in an enhanced ability to recognise bacterial infection. However, it is unclear how the two interleukins interact with the same signal transducer and lead to a different cellular response.11 ,16 ,18 With regard to the immune modulatory action of OM-85 BV, this finding could explain some of the IgG stimulation and enhanced host immune response.1-7 11-14Both factors act through the identical signalling transducer, gp130,16-18 28-30 and therefore exert similar cellular and immune modulatory effects.11-14 ,16 ,18 Both IL-6 and IL-11 are crucially involved in the early phase of host immune responses during infections. Interestingly, IL-6 and IL-11 reduced the infection dependent death rate of BALB/By mice treated withd-galactosamine plus Staphylococcus aureus enterotoxin B.11
With regard to the downregulating effect on IL-12 expression in lymphocytes achieved with preconditioned medium obtained from OM-85 BV stimulated fibroblasts, it is noteworthy that administration of recombinant IL-11 suppressed IL-12 production in lymphocytes.26 The induction of IL-11 by OM-85 BV could therefore explain its observed inhibitory effect on the expression of IL-12 in peripheral blood lymphocytes.
There is increasing evidence to suggest that IL-6 and IL-11 should be regarded as cell and organ protective factors rather than pro-inflammatory. The immunomodulatory effects ascribed to IL-6 and IL-11 seem to be mediated via their interaction with gp130.11 ,16 ,18 ,28-30 Activation of gp130, the only known signal transducer for both interleukins, requires binding of the respective cytokine to its corresponding specific receptor and subsequent complex formation with the signal transducer gp130.11 ,16 ,18 ,28-30 However, regulation of gp130 by various factors including interleukins,28-30 growth factors,28 ,30 and glucocorticoids29 is controversial and may be caused by different cell types. In line with our findings, the expression of gp130 seems to be mainly constitutive and can be only marginally modulated by endogenous factors.28-30 Similar to our findings of OM-85 BV induced accumulation of gp130 protein, dexamethasone, a strong cell differentiation factor, induced membranous accumulation of gp130 but not a significant enhanced transcription in HepG2 cells.29Unfortunately the molecular biological pathway controlling the expression of gp130 is still unclear.
In summary, our findings on the coordinated OM-85 BV induced expression of IL-6 and IL-11, their corresponding receptors, and the signal transducer gp130 may explain some of the beneficial immune supportive effects of OM-85 BV observed in patients with pulmonary infections.1-7