Article Text

Abstract
BACKGROUND It has previously been shown that M1 cholinergic receptors are involved in the parenchymal response to inhaled methacholine in puppies using the M1 selective antagonist pirenzepine. Although M3 receptors are responsible for acetylcholine induced bronchoconstriction in isolated rat lung, the role of M1receptors has not been determined in the rat in vivo.
METHODS Anaesthetised, paralysed, open chested Brown Norway rats were mechanically ventilated and the femoral vein cannulated for intravenous injection of drugs. Low frequency forced oscillations were applied to measure lung input impedance (Zl) and computerised modelling enabled separation of Zl into airway and parenchymal components. Atropine (500 μg/kg iv) and pirenzepine (50, 100 or 200 μg/kg iv) were administered during steady state constriction generated by continuous inhalation (1 mg/ml) or intravenous (10 or 15 μg/kg/min) administration of methacholine.
RESULTS Continuous inhalation of methacholine produced a 185% increase in frequency dependent tissue resistance (G) which was effectively inhibited by atropine 500 μg/kg iv (p<0.01, n = 6). Pirenzepine (50, 100 or 200 μg/kg) had a minimal effect on the parenchymal response to inhaled methacholine. A 258% increase in airway resistance (Raw) was induced by continuous intravenous infusion of methacholine and this response was effectively abolished by pirenzepine (p<0.001, n = 5). Cutting the vagi in the cervical region did not alter baseline airway mechanics. Vagotomy did not affect lung responses to intravenous methacholine nor the ability of pirenzepine to reduce these responses.
CONCLUSIONS In the rat, M1-subtype receptors are functional in airways but not in the tissue.
- forced oscillation technique
- muscarinic blockade
- lung parenchyma
Statistics from Altmetric.com
Methacholine is commonly used as a challenge agent for the assessment of hyperresponsiveness in asthmatic subjects. It is a chemical analogue of acetylcholine and induces smooth muscle contraction by stimulating muscarinic cholinergic receptors located on the smooth muscle. In a number of animal studies the way in which methacholine alters lung function has been shown to depend on the route of delivery. Intravenous methacholine acts mainly on the airway producing an increase in airway resistance whereas inhaled methacholine alters the mechanical properties of both the airways and the lung tissues.1-3 One possible explanation for these findings is that different receptors are involved.
Muscarinic cholinergic receptors exist in at least four major subtypes that can be demonstrated pharmacologically.4 Multiple subtypes have, however, been identified biochemically in the lung and binding studies have shown the presence of M1, M2 and M3 subtypes.5 ,6 A species difference in distribution of muscarinic receptors of the lung has been established. In isolated rat lung, stimulation of the M3receptor subtype is responsible for acetylcholine induced bronchoconstriction.7 However, previous experiments in our laboratory using alveolar capsules in puppies have shown that the peripheral lung responses to inhaled methacholine were antagonised by the M1 selective blocker pirenzepine.8
Recent adaptations of the low frequency forced oscillatory technique in rodents have enabled us to partition lung function changes into airway and lung tissue components.1 Peripheral lung is a complex anatomical structure comprising bronchial wall (approximately 5%), vascular smooth muscle (8%), and alveolar ducts and walls (>86%) in rats.9 The present experiments were performed to investigate whether the peripheral lung responses to inhaled methacholine resulted from stimulation of muscarinic cholinergic receptors. Based on our previous findings in puppies, the role of M1 receptors in airway and tissue responses to methacholine was also investigated by using the M1 selective antagonist pirenzepine.10
Methods
ANIMAL PREPARATION
Experiments were performed using adult male Brown Norway rats weighing 280–320 g. The rats were anaesthetised by intramuscular injection of xylazine (12 mg/kg) and ketamine (40 mg/kg) and placed in the supine position. The trachea was exposed in the mid cervical region and cannulated with a metal catheter (internal diameter 2 mm, length 1 cm) and the animals were mechanically ventilated (Harvard Rodent Ventilator 683, MA, USA) with a tidal volume of 9 ml/kg and a frequency of 90 breaths/minute. The femoral veins were cannulated with polyethylene tubing for injection of drugs. Paralysis was achieved with pancuronium bromide (0.2 mg/kg). The thorax was opened by midline thoracotomy with the positive end expiratory pressure set to 2.5 cm H2O and the ribs were widely retracted.
Maintenance doses of anaesthetic and muscle relaxant were administered intravenously every 45 minutes. The electrocardiogram and heart rate were monitored continuously using limb leads connected to a standard electrocardiograph (78342A, Hewlett Packard Co, Idaho, USA). Vagi were intact except in the third group of experiments where both right and left vagi were sectioned in the cervical region before baseline measurements and administration of intravenous methacholine and pirenzepine.
ESTIMATION OF AIRWAY AND PARENCHYMAL PARAMETERS
Lung input impedance (Zl) was measured using an adaptation of the low frequency forced oscillation technique in which pressure is measured at either end of a wave tube.1 The experimental set up is shown in fig 1. A three-way tap was used to switch the animal from the respirator to a loudspeaker-in-box system at end of expiration. The pressure in the box was set to 2.5 hPa to keep the transpulmonary pressure constant during measurements. The loudspeaker generated a small amplitude pseudorandom signal (15 non-integer multiples between 0.5 and 21 Hz) through a 120 cm long polyethylene wave tube with an internal diameter of 2 mm. Two identical pressure transducers (ICS model 33NA002D) were used to measure the lateral pressures at the loudspeaker (P1) and at the tracheal end (P2) of the wave tube. The P1 and P2 signals were low pass filtered (5th order Butterworth, 25 Hz corner frequency) and sampled with an analogue to digital board of an IBM compatible computer at a rate of 128 Hz. Fast Fourier transformation with time windows of four seconds and 95% overlapping was used to calculate the pressure transfer functions (P1/P2) from the six second long recordings. Zl was calculated as the load impedance of the wave tube using the following equation:

Schematic representation of the experimental set up. A loudspeaker is used to generate the low frequency forced oscillation signal which is delivered to the animal via a wave tube. Pressure is measured at each end of the wave tube and used to calculate the input impedance of the animal. The use of the wave tube obviated the need to measure flow in small animals. During measurements the ventilator is switched out of the circuit using a three-way tap.
Zl = Zo sinh(γL)/[(P1/P2)—cosh(γL)]
where L is the length, Zo is the characteristic impedance, and γ is the complex propagation wave number of the wave tube. The latter two parameters are determined by the geometrical data and the material constants of the tube wall and the air.
To separate the airway and parenchymal mechanics, a model containing a frequency independent airway resistance (Raw) and inertance (Iaw) in series with a constant phase tissue model including frequency dependent tissue resistance (G) and tissue elastance (H) was fitted to the Zl spectra by minimising the differences between the measured and modelled impedance values using the following equation:
Zl = Raw + jωIaw + (G—jH)/ωα
where j is the imaginary unit, ω is the angular frequency (2πf), and α, which is expressed as α = 2/π arctan (H/G), which is not an independent parameter. Impedance points coinciding with the heart rate and its harmonics were omitted from the model fitting since cardiac activity caused low signal-to-noise ratio at these frequency components. A schematic representation of this model is shown in fig2.
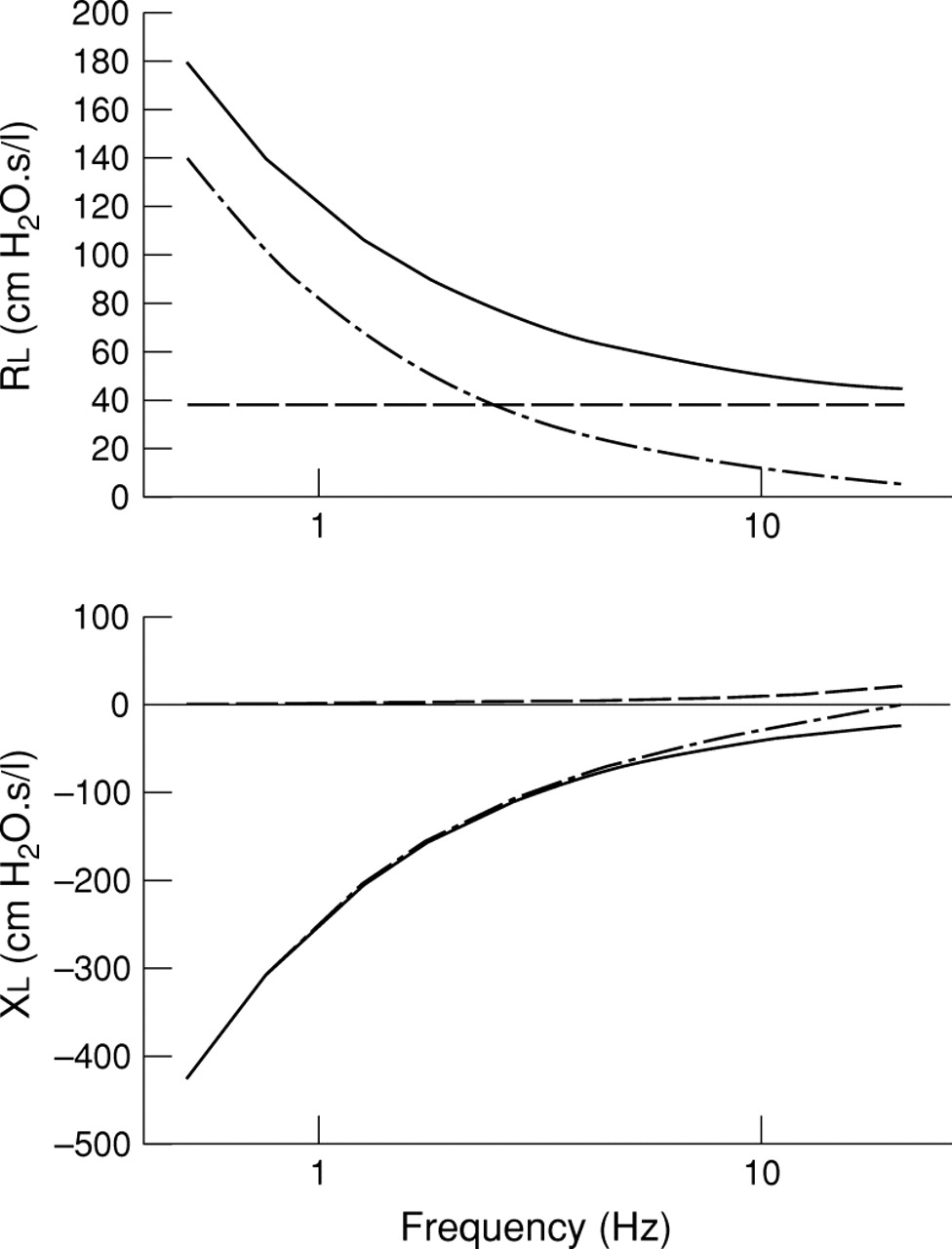
Schematic representation of the parameters derived from the constant phase model. The input impedance (Zl) is shown with solid lines. In the upper panel the real part of Zl (resistance) can be modelled as the combination of a frequency independent airway resistance (dashed line) and a frequency dependent tissue resistance (dash-dot line). In the lower panel the imaginary part of Zl (reactance) can be modelled as the combination of a frequency independent airway inertance (dashed line) and a frequency dependent tissue elastance (dash-dot line).
RESPONSES TO METHACHOLINE
To standardise volume history the lungs were hyperinflated by superimposing two inspiratory cycles before starting measurements. Following 15 minutes of mechanical ventilation, baseline respiratory mechanics were measured by recording 4–6 Zl spectra. These were ensemble averaged and the model fitted to the average spectrum. Methacholine was then continuously administered in a concentration sufficient to increase frequency dependent tissue resistance (G) by 50–100% at the steady state response.
INHALED METHACHOLINE
Methacholine was administered to the animal by inhalation at a concentration of 1 mg/ml using a jet nebuliser (LC PLUS, Pari-Werk GmbH, Germany) driven by compressed air (5 l/min) into the inspiratory port of the respirator. This nebuliser delivers particles approximately 5 μm in size (manufacturer’s specifications). Steady state constriction was obvious after 8–12 minutes.
INTRAVENOUS METHACHOLINE
Methacholine was administered intravenously by continuous infusion at a rate of 10–15 μg/kg/min (Stoelting 220VAC, USA), sufficient to increase airway resistance (Raw) by approximately 100%. Individual Zl curves were collected every two minutes until a plateau response was observed (approximately 15 minutes). These procedures resulted in a total methacholine administration time of 25–35 minutes.
RESPONSES TO MUSCARINIC ANTAGONIST
After the steady state response to methacholine had been established, the antagonist was administered by intravenous injection. After a period of two minutes individual Zl curves were collected also at two minute intervals. Atropine sulphate (500 μg/kg) was administered intravenously during the steady state response to inhaled methacholine. Pirenzepine was administered in cumulative doses of 50, 100 or 200 μg/kg at approximately 10–15 minute intervals during steady state response to either inhaled or intravenous methacholine.
EFFECTS OF CERVICAL VAGOTOMY
In animals with the vagi cut, the effect of cumulative administration of pirenzepine (50, 100, or 200 μg/kg) during steady state constriction induced by intravenous methacholine was investigated following baseline measurements. The infused concentration of methacholine used to approximately double Raw was 10 μg/kg/min in four rats and 15 μg/kg/min in one animal.
MATERIALS
Pirenzepine was kindly donated by Boehringer Mannheim GmbH (Mannheim, Germany); atropine sulphate and pancuronium bromide was purchased from Astra Pharmaceuticals (North Ryde, NSW, Australia); acetyl-β-methylcholine chloride (methacholine) was obtained from Sigma Chemical Company (St Louis, MO, USA); ketamine was purchased from Troy Laboratories (Smithfield, NSW, Australia); xylazine from Bayder (Pymble, NSW, Australia); and pentobarbitone sodium from Virbac (Peakhurst, NSW, Australia).
STATISTICAL ANALYSIS
Comparisons between groups were made on log transformed data using one way analysis of variance (ANOVA) with Student-Newman-Keuls correction for multiple comparisons. All results are expressed as mean (SE). p values of <0.05 were regarded as statistically significant.
Results
LUNG FUNCTION IN RESPONSE TO INHALED METHACHOLINE
Impedance spectra at baseline and after methacholine challenge are shown in fig 3. During steady state constriction induced by continuous inhalation of methacholine, frequency dependent tissue resistance (G) was increased by 185% above baseline (p<0.01, n = 6; fig 4). This increase in G was associated with a similar 142% increase in tissue elastance (H) (p<0.01, n = 6). Inhaled methacholine produced only minimal changes in airway resistance (Raw) and no change in airway inertance (Iaw) (fig 4).
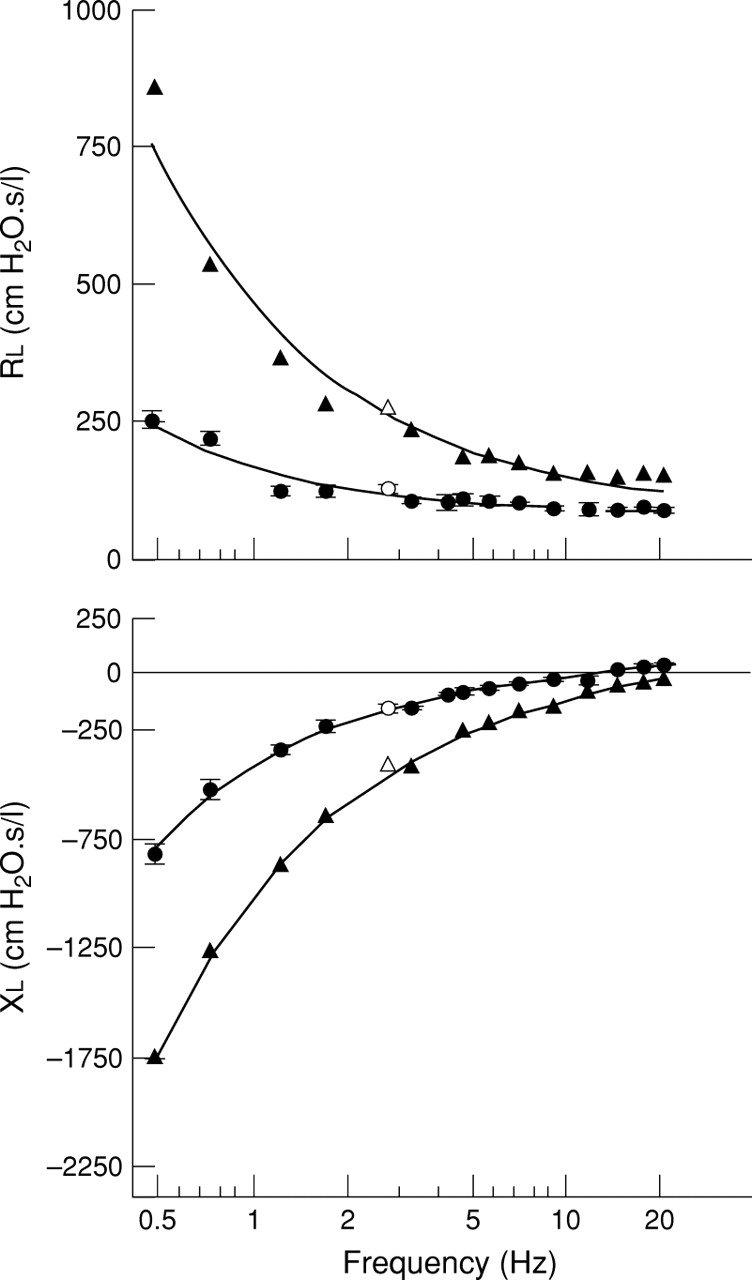
Real (lung resistance Rl) and imaginary (lung reactance Xl) parts of lung input impedance spectra (Zl) and the corresponding model fits in a single rat after inhalation of saline (•) and methacholine 5 mg/ml (▴). With saline, values are given as mean (SE) of 4–6 successive Zlrecordings. Following methacholine administration only the peak response is reported. Open symbols denote data points corrupted by cardiac noise and therefore omitted from the model fit.

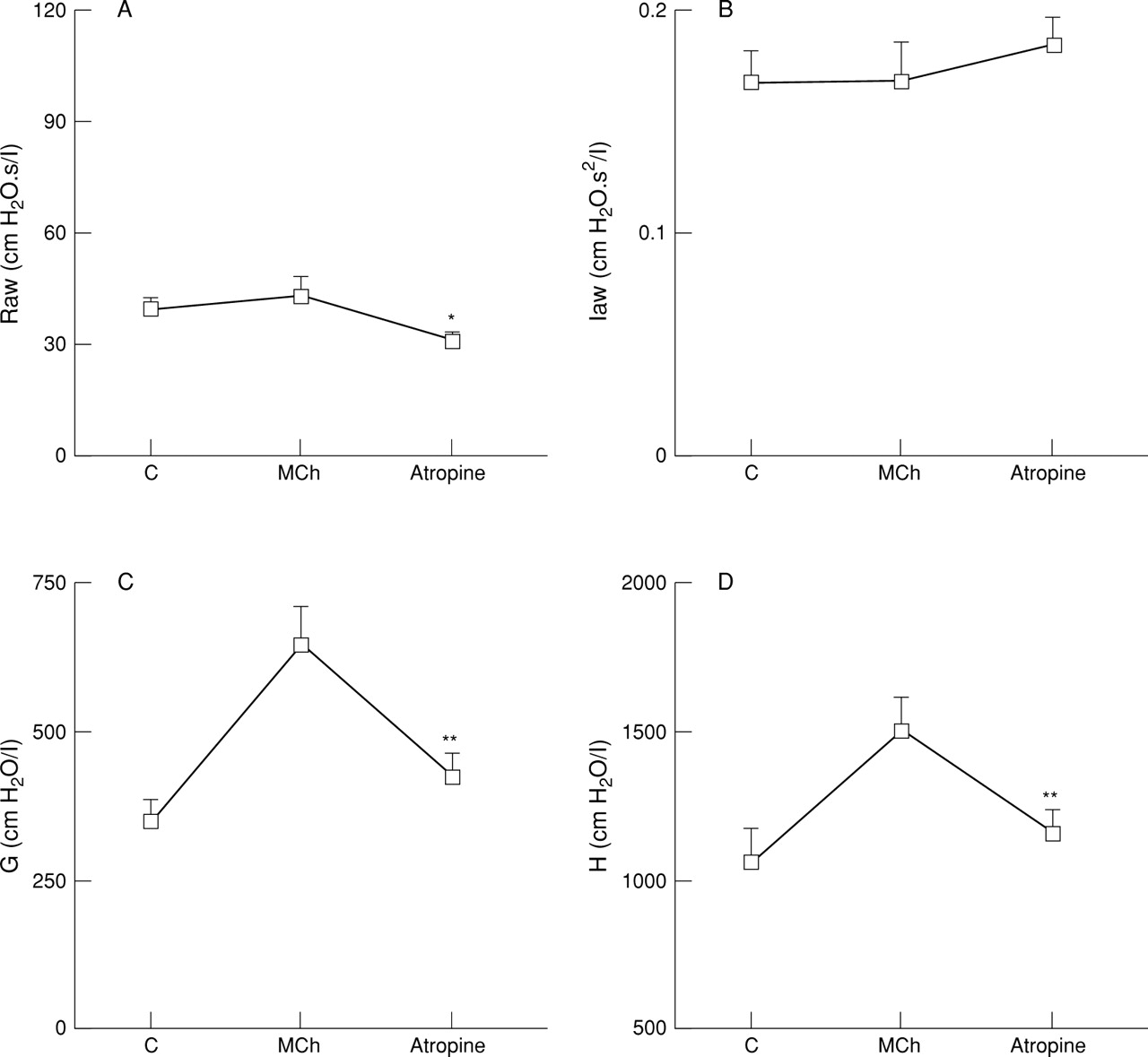
Effect of atropine sulphate (500 μg/kg iv) on (A) airway resistance (Raw), (B) airway inertance (Iaw), (C) frequency dependent tissue resistance (G), and (D) tissue elastance (H) during steady state response to inhaled methacholine (1 mg/ml) in anaesthetised rats with vagi intact. Results are given as mean (SE) of six animals. *p<0.05, **p<0.01 versus methacholine response.
EFFECT OF ATROPINE ON INHALED METHACHOLINE
Following intravenous injection of atropine (500 μg/kg iv) parenchymal responses to inhaled methacholine were inhibited, significantly reducing G by 62% (p<0.01, n = 6) and H by 77% (p<0.01, n = 6; fig 4). While G was still significantly above the baseline or control value after atropine treatment (p<0.001), tissue elastance (H) returned to the baseline level after atropine administration. Despite the lack of effect of methacholine on Raw, intravenous injection of atropine significantly reduced the baseline airway tone by 20% (p<0.05, n = 6). Little change was observed in Iaw with methacholine or atropine administration.
EFFECT OF PIRENZEPINE ON INHALED METHACHOLINE
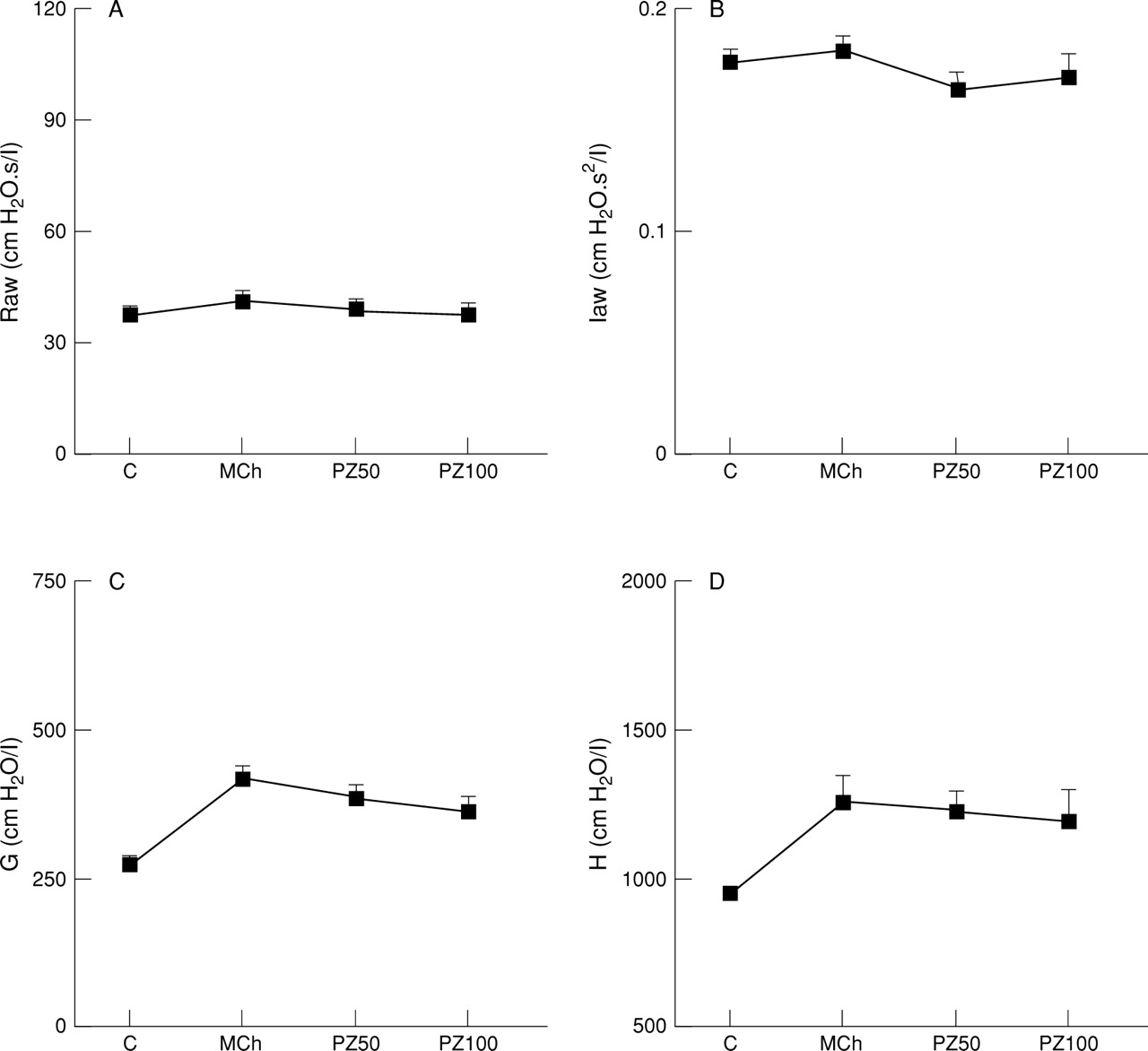
During steady state constriction induced by inhaled methacholine G was increased by 153% above baseline (p<0.01, n = 5; fig 5) and was associated with a corresponding 132% increase in tissue elastance (p<0.05, n = 5). Once again, no change in Raw or Iaw was evident with inhaled methacholine. Pirenzepine (50 μg/kg iv) produced no effect on parenchymal responses to methacholine (fig 5). Increasing the cumulative dose of pirenzepine to 100 μg/kg (fig 5) and 200 μg/kg (results not shown) produced no additional effect.

Effect of pirenzepine 50 μg/kg iv (PZ50) or 100 μg/kg iv (PZ100) on (A) airway resistance (Raw), (B) airway inertance (Iaw), (C) frequency dependent tissue resistance (G), and (D) tissue elastance (H) during steady state response to inhaled methacholine (1 mg/ml) in anaesthetised rats with vagi intact. Results are given as mean (SE) of five animals.
LUNG FUNCTION IN RESPONSE TO INTRAVENOUS METHACHOLINE: VAGI INTACT
The airway and tissue steady state responses to continuous intravenous infusion of mthacholine are shown in fig 6. Intravenous methacholine induced a marked and statistically significant increase in airway resistance of 258% above the baseline value (p<0.001, n = 6; fig 6A) and an increase of 165% in frequency dependent tissue resistance (p<0.01, n = 6; fig 6C). Intravenous administration of methacholine produced no effect on airway inertance (fig 6B) or tissue elastance (fig 6D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of pirenzepine 50 μg/kg iv (PZ50) or 100 μg/kg iv (PZ100) on (A) airway resistance (Raw), (B) airway inertance (Iaw), (C) frequency dependent tissue resistance (G), and (D) tissue elastance (H) during steady state response to intravenous methacholine (10 mg/kg/min) in anaesthetised rats with vagi intact (○, n = 5) or vagi cut (•, n = 5). Results are given as mean (SE).
EFFECT OF PIRENZEPINE ON INTRAVENOUS METHACHOLINE: VAGI INTACT
Intravenous injection of pirenzepine (50 μg/kg) resulted in a rapid decline in both the airway and tissue response to methacholine, reducing Raw by 76% (p<0.001, n = 6) and G by 73% (p<0.001, n = 6) from the plateau steady state methacholine response (fig 6). Iaw was significantly reduced by 21% below baseline (p<0.05, n = 6) and H remained unchanged (fig 6). The maximum effect on Raw and G was evident within five minutes after administration and was maintained over the period of 15 minutes in which the response was observed.
Increasing the dose of pirenzepine to 100 μg/kg resulted in further reduction in airway resistance to 97% from plateau methacholine response or 3% above baseline (p<0.05, n = 6), completely abolishing the airway response to methacholine. Similarly, G was further reduced to 77% of the methacholine plateau response (p<0.001, n = 6; fig 6). The response in G was still significantly higher than the baseline response (p<0.05). Increasing the pirenzepine dose to 200 μg/kg iv (results not shown) did not result in further attenuation of the parenchymal responses to intravenous methacholine.
LUNG FUNCTION IN RESPONSE TO INTRAVENOUS METHACHOLINE: VAGI CUT
Cervical vagotomy produced no effect on baseline lung function parameters when compared with animals with vagi intact (fig 6). In the airway compartment a small reduction in the baseline Raw (15%) in the rats with vagi cut failed to reach statistical significance. With the vagi cut, the airway response to intravenous methacholine was similar to that seen with the vagi intact (fig 6), with Raw increasing by 284% (p<0.05, n = 5) above baseline. The same pattern of response was also seen for G which increased by 191% from baseline (p<0.01, n = 5). In rats with cervical vagotomy only, minor but significant increases in Iaw (12%) and H (16%) were apparent after methacholine administration (fig 6).
EFFECT OF PIRENZEPINE ON INTRAVENOUS METHACHOLINE: VAGI CUT
Administration of pirenzepine in a dose of 50 μg/kg iv effectively reduced both the airway and the parenchymal response to intravenous methacholine, reducing Raw by 78% (p<0.05, n = 5) and G by 67% (p<0.05, n = 5) from the plateau steady state methacholine response. Pirenzepine in a dose of 50 μg/kg iv significantly reduced the baseline response in Iaw (p<0.01, n = 5) and H remained unchanged (fig 6). Increasing the dose of pirenzepine to 100 μg/kg further reduced Raw to 94% of the plateau response (p<0.001, n = 5) with no additional significant effect on G (fig 6). Pirenzepine 200 μg/kg (results not shown) did not produce any further effect. The pattern and time course of the pirenzepine response in animals with vagi cut was similar to the responses in animals with vagi intact.
Discussion
The results of these experiments in anaesthetised Brown Norway rats show that prolonged inhalation of methacholine results in a predominantly parenchymal (or tissue) constrictor response which is effectively reversed by a maximum dose of atropine sulphate but not by pirenzepine. In contrast, intravenous methacholine produced more marked changes in airway resistance which were reversed by pirenzepine.
Our adaptation of the low frequency forced oscillation technique allows lung mechanics to be partitioned into airway and parenchymal components by fitting the constant phase model to the impedance spectra. The airways through which gas moves by bulk flow will be included in the airway component, whereas airways through which gas moves by diffusion are likely to be included in the parenchymal component. It is therefore most likely that the parenchymal response is the result of contraction of smooth muscle in respiratory bronchioles and alveolar ducts and of contractile elements in the alveolar septa. These may be the contractile elements described by Kapanci et al.11 Vascular smooth muscle is present in the lung parenchyma but may be expected to relax in response to methacholine.12 Both atropine and cervical vagotomy tended to decrease Raw. Due to the variability, this reached statistical significance only for atropine. The lack of effect on baseline parenchymal measurements is consistent with cholinergic innervation being most dense in the large airways.13 Methacholine responses on airways were not altered by removing vagal tone.
Atropine is a non-selective muscarinic antagonist with very similar potency on all muscarinic receptor subtypes.14 It was effective in the present experiments in reducing the parenchymal responses to inhaled methacholine, confirming the role of muscarinic receptors in producing this response. A significant reduction in Raw was also apparent. Pirenzepine demonstrates relative selectivity for M1 subtypes.14 ,15 In the present model pirenzepine inhibited the airway responses to intravenously administered methacholine but, unlike atropine, did not reduce the tissue responses to inhaled methacholine. The lowest dose of pirenzepine (50 μg/kg iv) was approximately the ED50dose, as reported by other workers measuring inhibition of the increased blood pressure with the M1 selective agonist McN-A-343 in anaesthetised rats.15 In the present experiments pirenzepine reduced the airway response to intravenously infused methacholine by 76% and 78% in rats with vagi intact and severed, respectively. Pirenzepine (100 μg/kg iv) had an almost maximal effect on the Raw to intravenous methacholine and yet, at this dose, there was no significant reduction in tissue responses to inhaled methacholine. It is therefore likely that pirenzepine is not antagonising M3 receptors at these concentrations. As pirenzepine has a selectivity profile of M1 > M4, M3 > M2,14 ,15 it is also unlikely that it is acting on M2 receptors and one must conclude that we are observing an M1 effect on Raw. Few M1 subtype muscarinic receptors have been identified in rat lung using binding studies,5 ,6 and only M2 and M3 receptors were evident in the trachea and bronchi.6 Up to 91% of muscarinic receptor protein in rat lung is of the M2 subtype with M3 receptor protein making up most of the remainder.8 The apparently small number of M3 receptors has, however, been reported to dominate muscarinic responses.7 It is therefore possible that a small number of M1 receptors will still be sufficient to produce a physiological response.
Pirenzepine reversed the changes in both Raw and G when constriction was induced by intravenous methacholine. The increase in G with methacholine was not, however, associated with an increase in H. Lutchen et al 16 investigated this phenomenon using ventilation with gas mixtures of differing viscosities and densities. They explained the increase in G as resulting from heterogeneous airway constriction. With this background we can interpret our results as follows. Pirenzepine competitively reverses the highly inhomogeneous constriction of the bronchial tree present following intravenous methacholine, therefore decreasing the inhomogeneities present in the periphery and producing a decrease in G. Taking into account the pirenzepine results when lung constrictor tone is induced either by aerosolised or intravenously administered methacholine, our data suggest that pirenzepine acts only on the airways since a decrease in G was not evident when a purely parenchymal response to inhaled methacholine was generated.
M1 receptors are understood to be associated with neuronal tissue to facilitate parasympathetic ganglionic transmission17 which, with the generalisation that autonomic innervation decreases down the respiratory tree,13 would agree with the purely airway effects observed in our experiments. They were also present in human alveolar walls in human lungs but not in the guinea pig.18 The ability of pirenzepine to reverse the airway responses to methacholine could be explained if methacholine resulted in a positive feedback, via ganglionic M1 receptors, resulting in increased release of acetylcholine from cholinergic nerves. However, the fact that the same pattern of response was seen after bilateral cervical vagotomy makes this explanation unlikely. While caution must be exercised when extrapolating from the results of animal studies to humans, clinical studies have shown that pirenzepine improves lung function in both healthy subjects19 and asthmatics.20
In conclusion, the present experiments in anaesthetised rats show that, using lung impedance measurements, the non-selective muscarinic antagonist atropine reverses the increase in parenchymal resistance produced by prolonged inhalation of methacholine. The M1selective muscarinic antagonist pirenzepine had no significant effect on parenchymal responses to inhaled methacholine and reversed airway responses to intravenously administered methacholine. Cutting the cervical vagi did not alter resting lung function or responses to methacholine and pirenzepine. It therefore appears that, in the rat, M1 subtype receptors are functionally present in the airways but could not be demonstrated in the parenchyma.
Acknowledgments
This study was supported by a grant from the National Health and Medical Research Council, Australia and an Australian Lung Foundation, Smith Kline and Beecham Award. M K Tulić is holding an Asthma Foundation of Western Australia Postgraduate PhD Scholarship.