Article Text

Abstract
Background Recent evidence indicates that human regulatory T cells (Tregs) are composed of three distinct subpopulations: CD25++ CD45RA+ resting Tregs (rTregs), CD25+++ CD45RA− activated Tregs (aTregs), which are suppressive, and CD25++ CD45RA− cytokine-secreting (Fr III) cells with pro-inflammatory capacity.
Objectives To evaluate the dynamic changes in circulating and pulmonary Treg subpopulations in smokers and patients with chronic obstructive pulmonary disease (COPD), and to explore their potential roles in COPD pathogenesis.
Methods Blood samples were obtained from 57 never-smokers, 32 smokers with normal lung function and 66 patients with COPD. Bronchoalveolar lavage (BAL) samples were taken from 12 never-smokers, 12 smokers and 18 patients with COPD. The proportions of Treg subpopulations and activated CD8 T cells were evaluated using flow cytometry.
Results In peripheral blood, increased proportions of rTregs, aTregs and Fr III cells were found in smokers compared with never-smokers, whereas patients with COPD showed decreased rTregs and aTregs, and significantly increased Fr III cells compared with smokers. The changes in Treg subpopulations, with an overall decrease in the (aTreg+rTreg):(Fr III) ratio, indicated that immune homeostasis favoured inflammation and correlated with enhanced CD8 T-cell activation (r=−0.399, p<0.001) and forced expiratory volume in 1 s (FEV1) % predicted value (r=0.435, p<0.001).The BAL (aTreg+rTreg):(Fr III) ratios displayed more robust correlations with FEV1% predicted value (r=0.741, p<0.01) and activation of effector T cells (r=−0.763, p<0.001).
Conclusions The imbalance between the anti-inflammatory subsets (aTreg+rTreg) and the pro-inflammatory subset (Fr III) of Tregs may play an important role in COPD progression.
- COPD AU Mechanisms
- Lymphocyte Biology
- Bronchoscopy
Statistics from Altmetric.com
Key messages
What is the key question?
-
Are phenotypically and functionally different subpopulations of regulatory T cells (Tregs) varied in chronic obstructive pulmonary disease (COPD) and potentially involved in the progression of inflammation?
What is the bottom line?
-
The imbalance between the anti-inflammatory subsets and the pro-inflammatory subset of Tregs correlates with loss of lung function and immune activation in patients with COPD.
Why read on?
-
This study provides evidence to support disturbed homeostasis of Treg subpopulations as a potential mechanism for persistent inflammation mediated by CD8 cells in COPD.
Introduction
The principal feature of chronic obstructive pulmonary disease (COPD) is abnormal inflammatory response of the lung to inhaled noxious gases or particles, especially to tobacco smoke.1 Accumulating evidence indicates that chronic inflammation and adaptive immunity play important roles in the development and progression of COPD.2–4 The inflammation in COPD extends beyond the lungs and includes regional lymph nodes and the systemic circulation.5–7 However, the regulation of inflammation in patients with COPD has not yet been fully elucidated.
T lymphocytes are believed to be key cells in regulating airway inflammation in COPD.7 Activated T cells, especially activated CD8 T cells, can cause a variety of the tissue injuries that typify COPD by direct cytopathic effects, elaboration of diverse pro-inflammatory and deleterious mediators, and/or recruitment and activation of other immune and parenchymal effector cells.8 ,9 A number of studies have demonstrated that activation of T cells is tightly controlled by active mechanisms and negative regulatory mechanisms. In the past decade, CD4+ CD25+ Foxp3+ T regulatory cells (Tregs), a critical subset of T cells that use a variety of mechanisms to suppress the immune response, have gained considerable attention,10–14 and Treg abnormalities have been described in many chronic inflammatory and autoimmune disorders.10 ,11 Recently, accumulating evidence indicates that patients with COPD exhibit many of the characteristics of a classical autoimmune response.15 ,16 For these reasons, several studies have evaluated the role of Tregs in adaptive immunity of COPD. In patients with COPD, Treg proportions fluctuate in bronchoalveolar lavage (BAL) or in different parts of pulmonary tissue.17–20 However, contrary to the current notion of systemic inflammation, the frequencies of Tregs in peripheral blood do not differ significantly among patients with COPD, healthy smokers and never-smokers.17–20
In most previous studies, Tregs have only been considered as a whole population. However, recent studies have demonstrated that human CD4+ CD25+ Foxp3+ Tregs are heterogeneous and comprise three distinct subpopulations, each with a precise phenotype and function. Apart from resting Tregs (rTregs, CD25++ CD45RA+) and activated Tregs (aTregs, CD25+++ CD45RA−), which are suppressive in vitro, human CD4+ CD25+ Foxp3+ Tregs also possess CD25++ CD45RA− cytokine-secreting T cells (Fr III) with pro-inflammatory capacity.21 This population lacks suppressive activity or has limited suppressive activity, but can produce interleukin (IL)-2, IL-17 and interferon (IFN)-γ,21–25 which are important for T-cell activation and pulmonary inflammation.26 ,27 Recent studies revealed the clinical relevance of subpopulations of Tregs in patients with autoimmune diseases,21 HIV infection22 and diabetes.23
The aim of this study was to explore whether there were abnormal distributions in the suppressive and/or the pro-inflammatory subpopulations of Tregs in the blood and the lung of patients with COPD and smokers by using this new identification strategy of Foxp3+ T-cell subsets, and whether the frequency of different Treg subsets was correlated with effector T-cell (CD8) activation and disease severity as defined by forced expiratory volume in 1 s (FEV1). The findings should provide new insights into the potential mechanisms underlying persistent inflammation and adaptive immunity in COPD.
Materials and methods
Study subjects
Sixty-six patients with COPD and 34 smokers with normal lung function were recruited in Beijing Tongren Hospital, Capital Medical University, China. Fifty-seven never-smokers with normal lung function were also recruited as controls. Written informed consent was obtained and the local research ethics committee approved this study (TRECKT 2008-14). Subject baseline characteristics are summarised in table 1. A total of 112 patients with COPD were consecutively invited but 46 declined.
Demographics and spirometry of all participants
Subjects with a smoking history of ≥20 pack-years and normal lung function (FEV1>80% predicted) were categorised as smokers. COPD diagnosis was established according to the definition supplied by the Global Initiative for Chronic Obstructive Lung Disease (GOLD) guidelines.28 Patients with COPD had FEV1/forced vital capacity ratio<70% and FEV1<80% of the predicted value post bronchodilators. All patients with COPD were clinically stable and had not experienced any exacerbations for ≥3 months preceding inclusion in the study. Participants with neoplasm, interstitial lung disease, autoimmune disease, diabetes, infection or other immune-related diseases were excluded.
BAL fluid samples were obtained from 18 patients with COPD, 12 smokers and 12 never-smokers. The reasons for bronchoscopy were haemoptysis (COPD, n=9 of 18; smokers, n=5 of 12; never-smokers, n=7 of 12); unexplained shortness of breath or weight loss (COPD, n=5 of 18; smokers, n=3 of 12; never-smokers, n=3 of 12); and investigation of abnormal chest radiograph findings (COPD, n=4 of 18; smokers, n=4 of 12; never-smokers, n=2 of 12).
Cell collection
Peripheral blood samples were collected in ethylenediaminetetraacetic acid treated tubes from each subject and were processed to measure peripheral blood mononuclear cells (PBMCs) for flow cytometry setup procedures. Blood samples were layered onto Ficoll-Paque Plus (Amersham Biosciences, Amersham, Bucks, UK), centrifuged (400 g for 20 min at 21°C), and PBMCs were harvested. Cells were washed once in divalent cation-free Hanks balanced salt solution at 300 g for 5 min at 4°C. PBMCs were resuspended and viable counts obtained.
BAL was performed and processed as previously described.29 Bronchoscopy was performed with a flexible fibreoptic bronchoscope (Olympus, Tokyo, Japan) under topical lidocaine. Four separate 50 mL aliquots of sterile 0.9% saline solution were instilled in one pulmonary segment of a right middle lobe, or a lobe not affected by radiographic or endobronchial abnormalities. The fluid recovered was filtered, washed twice in phosphate-buffered saline and resuspended at106 cells/ml in RPMI-1640 medium at 4°C. BAL fluid and peripheral blood samples were processed immediately after attainment.
Flow cytometry
Freshly obtained human PBMC and BAL samples were stained as previously described21 with anti-hCD4-FITC (e-Bioscience, San Diego, California, USA), anti-hCD25-PE-Cy5 (BD Biosciences, San Jose, California, USA), anti-hCD45RA-APC (BD Biosciences). Anti-hCD8-FITC, anti-hCD28-APC, anti-hCD38-APC, anti-hHLA-DR-PE and anti-hCD69-FITC (BD Biosciences) antibodies were used for surface marker staining of each subpopulations. Treg subpopulations were gated as shown in online supplementary figure S1. Details of the cell counting, intracellular staining, cell culture and in vitro suppression assay are provided in the online supplementary appendix.
Data analysis
Group data were expressed as mean and SEM or as median and IQR when appropriate. For data not distributed normally, comparisons between three groups were made using a one-way Kruskal–Wallis test (p<0.05 was considered statistically significant). If this test indicated significance, the Mann–Whitney test was used for post hoc analysis for comparison between two groups, with corrections of p values according to Bonferroni (p<0.017 was considered statistically significant). Correlation was assessed by calculating Spearman's rank correlation coefficient. Here, p<0.05 was considered statistically significant. Statistical analysis was performed using SPSS for Windows V.16.0 (Chicago, Illinois, USA).
Results
Demographic characteristics of study population
The characteristics of the patients with COPD, smokers and never-smokers with normal lung function are summarised in table 1. There was no difference between the groups in terms of age. The unequal sex ratio was mostly due to the much higher prevalence of COPD in men than in women in China.30 There was no significant difference in the smoking history of patients with COPD and smokers. Table 2 shows the main clinical and functional characteristics of participants who underwent bronchoscopy. The demographic characteristics of this study population were consistent with those who did not receive bronchoscopy.
Demographics and spirometry of participants who underwent bronchoscopy
Dissection of Foxp3+ cells into three subsets with different function and phenotypic characteristics
As described previously,21 CD4+ Foxp3+ T cells were separated into three subpopulations by the expression of CD25 and CD45RA: rTregs (CD45RA+ CD25++, Fr I), aTregs (CD45RA− CD25+++, Fr II) and cytokine-secreting subset (CD45RA− CD25++, Fr III) (figure 1A). As shown in figure 1A, the degrees of FoxP3 expression in these fractions were proportional to CD25 expression. Moreover, rTregs expressed high levels of CD31, a marker for primary thymic emigrants, but expression was lost during their post-thymic peripheral expansion, as reported previously.21 ,31 Compared with rTregs, aTregs showed lower CD31 expression but higher expression of Foxp3, cytotoxic T-lymphocyte antigen 4 (CTLA-4) and HLA-DR, and Ki-67, a nuclear protein expressed in proliferating cells (figure 1B). These data suggest that these cells are a proliferating, activated fraction of Tregs. Although Fr III cells expressed intermediate levels of Foxp3, decreased levels of CTLA-4, HLA-DR and Ki-67 were observed. Meanwhile, a higher amount of IL-17 and IFN-γ were detected in response to phorbol-12-myristate-13-acetate (PMA) and calcium ionomycin stimulation (figure 1C), which confirmed that this subpopulation represented previously described pro-inflammatory cytokine-secreting cells.21
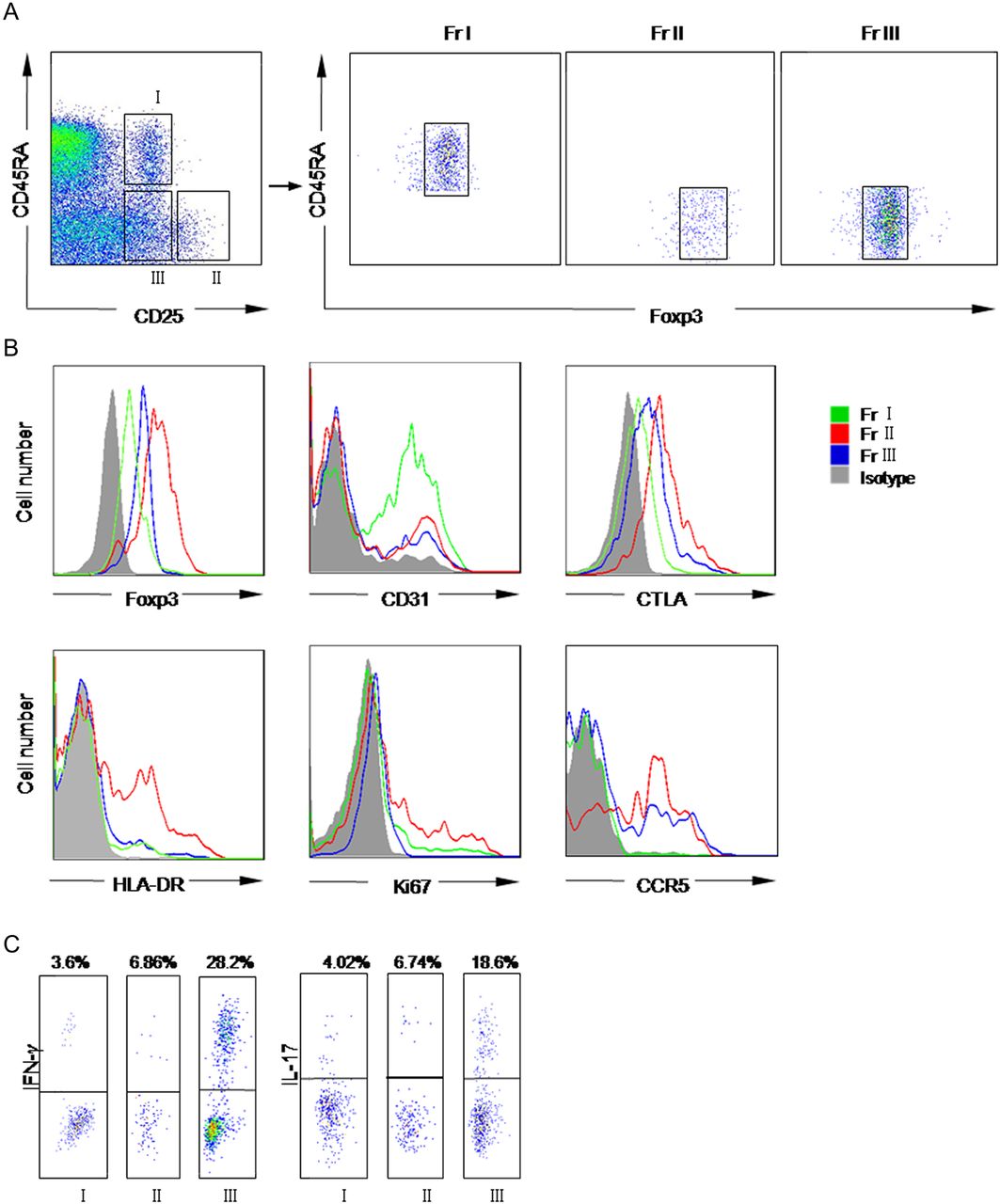
Dissection of CD4+ Foxp3+ T cells into three subsets by cell surface molecules and intracellular protein. (A) Three subsets of CD4 T cells defined by the expression of CD25, CD45RA (left panel). Right panels showed the FoxP3 expression in each subpopulation. (B) Each subpopulation expressed different amounts of Foxp3, cytotoxic T-lymphocyte antigen 4 (CTLA-4), CD31, HLA-DR, CD95 and Ki67. Fr I is represented by green lines, Fr II by red lines, Fr III by blue lines and isotype by solid grey profiles.(C) Flow cytometry of the production of interleukin 17 (IL-17) and interferon γ (IFN-γ) by three subsets after stimulation with phorbol-12-myristate-13-acetate (PMA) and ionomycin for 5 h. Percentages of cytokine-secreting cells are shown. Data are representative of five independent experiments. Fr I, CD25++ CD45RA+ resting T-regulatory cells; Fr II, CD25+++ CD45RA− activated T-regulatory cells; Fr III, CD25++ CD45RA− cytokine-secreting cells.
Varied subpopulation frequencies of circulating Tregs in patients with COPD, smokers and never-smokers
We first investigated the frequencies of Tregs in peripheral blood of different groups using the classic definition.32 In line with previous studies,17 ,18 Treg identification using CD4+ CD25+ Foxp3+ showed no difference in the circulating Treg number in patients with COPD (median 8.69%) compared with smokers (median 8.23%, p=0.45) and never-smokers (median 7.95%, p=0.16). However, the proportion of rTreg (Fr I), aTreg (Fr II) and cytokine-secreting (Fr III) fractions among CD4 T cells increased slightly in smokers compared with never-smokers (median, Fr I: 5.18% vs 3.25%, p<0.001; FrII: 2.01% vs 1.70%, p<0.01; Fr III: 7.37% vs 4.78%, p<0.001, for smokers and never-smokers, respectively). This suggested that tobacco smoke resulted in anti-inflammatory and inflammatory responses. Strikingly, in comparison to smokers, patients with COPD showed significantly increased proportions of Fr III cells (median 10.81%, p<0.001) but decreased proportions of rTregs (median 2.16%, p<0.0001) and aTregs (median 1.07%, p<0.0001; figure 2A–C). The number of each subset per microlitre of peripheral blood showed the same trend of variation as the proportion of each subset in different groups (figure 2D–F). Varied subpopulation frequencies suggested that the balance between immunosuppressive subsets and inflammatory subsets in CD4+ Foxp3+ T cells shifted towards inflammation in patients with COPD.

Variations in Foxp3+ cell subpopulations under physiological and disease conditions. (A–C) The percentages of each circulating T-regulatory subset among CD4 T cells in never-smokers, smokers and patients with chronic obstructive pulmonary disease (COPD). (D–F) Number of each Foxp3+ subset per microlitre of peripheral blood in never-smokers, smokers and patients with COPD. (G) Representative data of flow cytometry profiles are shown. Horizontal lines indicate median values. A p value<0.017 was considered statistically significant. Fr I, CD25++ CD45RA+ resting T-regulatory cells (rTreg); Fr II, CD25+++ CD45RA− activated T-regulatory cells (aTreg); Fr III, CD25++ CD45RA− cytokine-secreting cells.
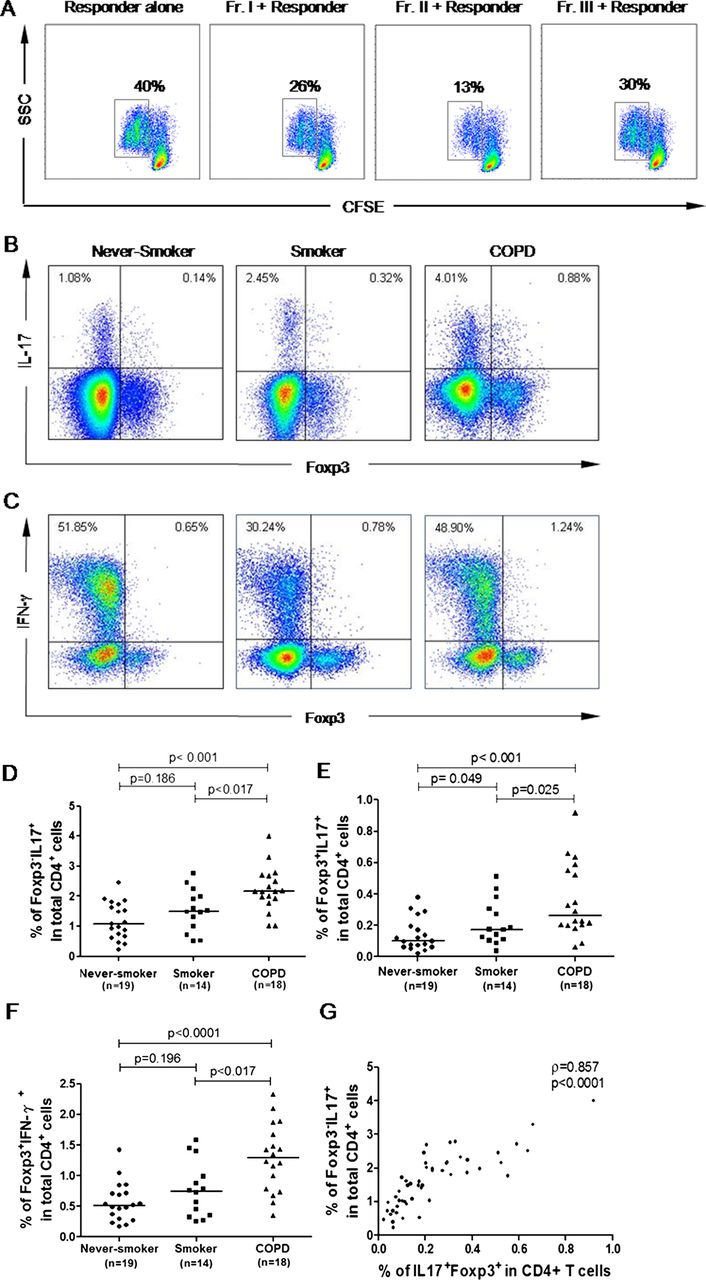
Next, we performed in vitro experiments to test the immune function of each population in patients with COPD. As shown in figure 3A, both rTregs and aTregs were functionally suppressive, while FrIII cells showed moderate suppressive capacity and could produce IL-17 and IFN-γ in response to PMA/ionomycin (figure 3B,C). In accordance with the phenotype results, compared with never-smokers, the numbers of Foxp3+ IL-17-secreting FrIII cells and Foxp3+IFN-γ-secreting FrIII cells were slightly increased in smokers, but dramatically increased in patients with COPD (figure 3E, F). More importantly, the number of Foxp3+ IL-17-secreting FrIII cells strongly correlated with T helper 17 (Th17) cells (Foxp3−IL17+) (r=0.818, p<0.001; figure 3G).
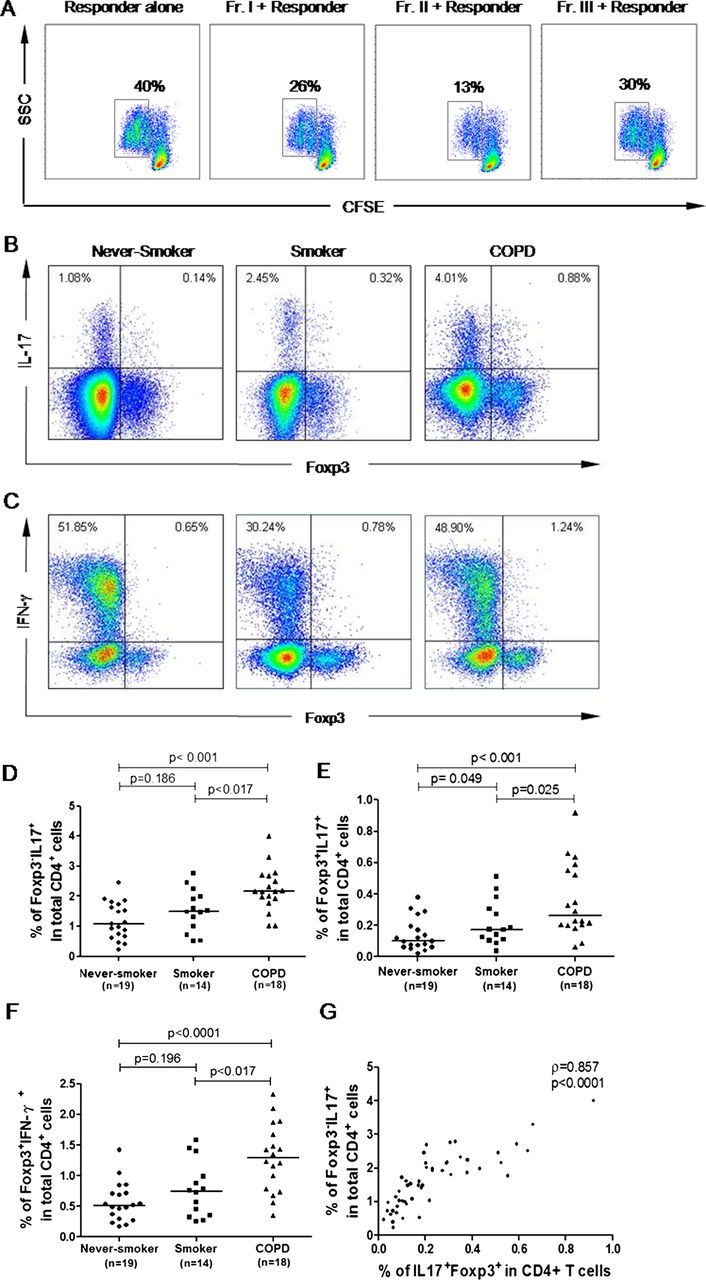
In vitro functional activity of the Foxp3+ cell subpopulation in patients with chronic obstructive pulmonary disease (COPD). (A) Suppression of T-cell proliferation by each subpopulation was assessed by carboxyfluorescein succinimidyl ester (CFSE) staining. Percentages of dividing cells were indicated. Data are representative of five independent experiments. (B and C) Intracellular staining of cytokines. Representative staining of intracellular Foxp3 versus interleukin 17 (IL-17) or interferon γ (IFN-γ) in never-smokers, smokers and patients with COPD after 5 h of stimulation with phorbol-12-myristate-13-acetate (PMA)/ionomycin and GolgiStop. Data are representative of 51 independent experiments. (D–F) Frequencies of Foxp3− IL-17+ (D), Foxp3+ IL17+ (E) and Foxp3+ IFN-γ (F) among CD4 T cells in three groups. A p value<0.017 is considered statistically significant. The top bar in (D–F) is for the difference between the never-smokers and patients with COPD. (G) Proportions of IL-17-secreting T-regulatory cells correlated with T helper 17 cells (Foxp3− IL17+). A p value<0.05 was considered statistically significant. Fr I, CD25++ CD45RA+ resting T-regulatory cells; Fr II, CD25+++ CD45RA− activated T-regulatory cells; Fr III, CD25++ CD45RA− cytokine-secreting cells.
Disturbed homeostasis among subpopulations of circulating Tregs correlated with effector T-cell activation status and disease severity
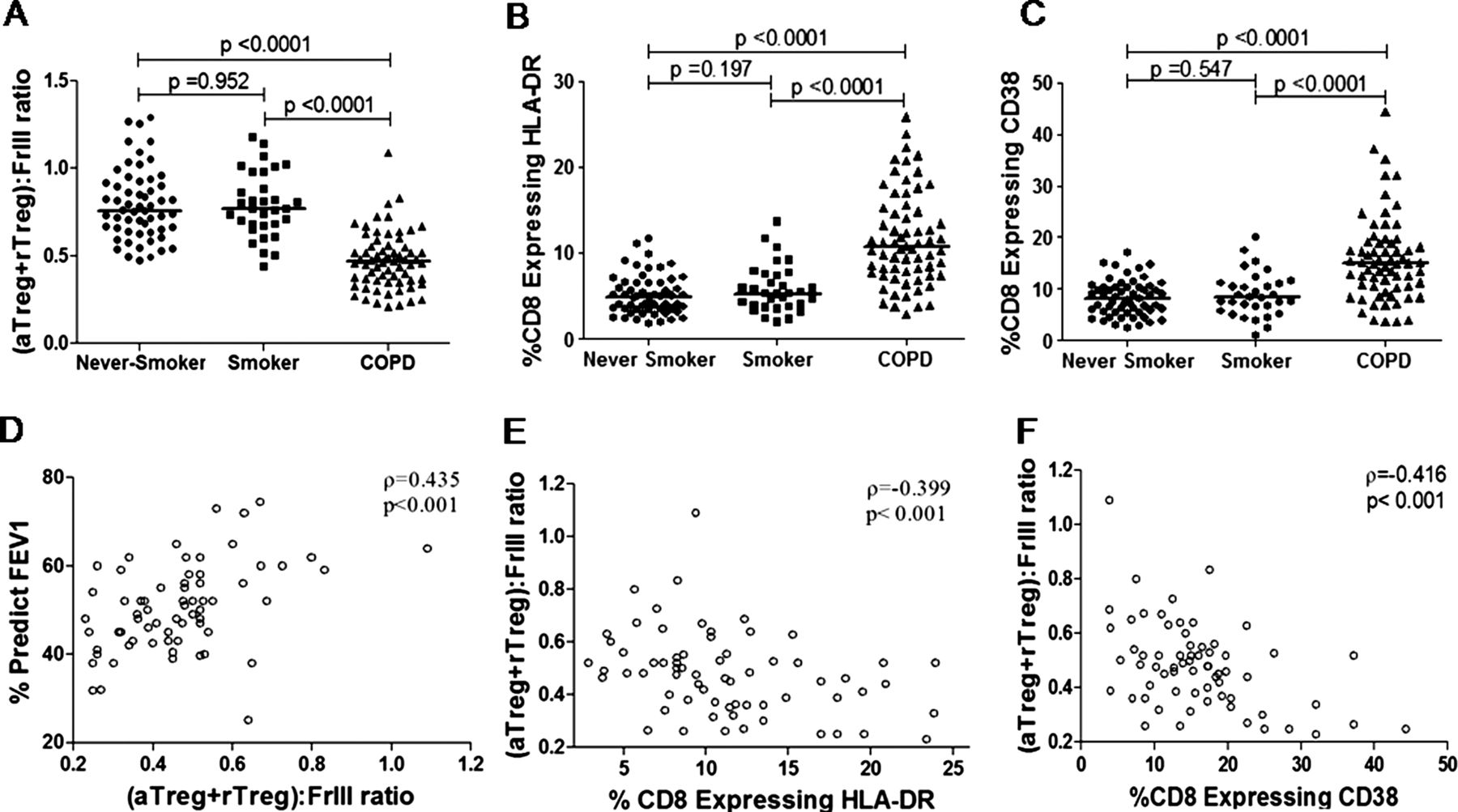
To illustrate further the balance between the suppressive and pro-inflammatory subpopulations, we calculated the ratio of (aTreg+rTreg):(Fr III). As expected, the (aTreg+rTreg):(Fr III) ratio was significantly decreased in patients with COPD (median 0.47) compared with smokers (median 0.77; p<0.0001) and never-smokers (median 0.75; p<0.0001) (figure 4A). No difference was found in the (aTreg+rTreg):(Fr III) ratio between the patients receiving and those not receiving inhaled corticosteroids (see online supplementary figure S2), which indicated that the differences in Treg subpopulations among never-smokers, smokers and patients with COPD were not due to corticosteroids. However, a definite conclusion could only be made by a prospective study to explore specifically the effect of corticosteroids on Tregs.

The imbalance between the suppressive and pro-inflammatory subpopulation correlated with disease severity and activation of CD8 T cells in the circulation. The (aTreg+rTreg):(Fr III) ratio was significantly decreased in patients with chronic obstructive pulmonary disease (COPD) compared with smokers and never-smokers (A); CD8 cells coexpressing HLA-DR (B) or CD38 (C) were increased in patients with COPD. A p value<0.017 was considered significant. The (aTreg+rTreg):(Fr III) ratio correlated with forced expiratory volume in 1 s (FEV1) predicted values (D) and CD8 T-lymphocyte activation status (E,F) in patients with COPD. A p value<0.05 was considered statistically significant. Horizontal lines indicate median values. Fr I, CD25++ CD45RA+ resting T-regulatory cells (rTreg); Fr II, CD25+++ CD45RA− activated T-regulatory cells (aTreg); Fr III, CD25++ CD45RA− cytokine-secreting cells.
It has been previously demonstrated that patients with COPD exhibited enhanced CD8 T-lymphocyte activation, which was correlated with the disease severity as measured by FEV1.33 Moreover, it has been shown that CD8 T-cell activation could be suppressed by Tregs10 and promoted by IL-17.34 ,35 The decreased (aTreg+rTreg):(Fr III) ratio in patients with COPD prompted us to test whether the disturbed Treg homeostasis had any influence on CD8 T-cell activation and diseases severity. The expression of CD28 (a costimulatory molecule), CD69 (an early marker transiently expressed on activated T cells), CD38 and HLA-DR (more general markers of T-cell activation) on CD8 T cells was evaluated. Similar to a previous study,33 patients with COPD showed elevated frequencies of CD8+HLA-DR+ (median 10.72%, 5.33% and 4.98% for patients with COPD, smokers and never-smokers, respectively, p<0.001) and CD8+ CD38+ cells (median 14.95%, 8.40% and 8.24% for patients with COPD, smokers and never-smokers, respectively, p<0.001) (figure 4B,C). However, expression of CD28 and CD69 in CD8 T lymphocytes did not differ significantly between the groups (data not shown).
In line with previous reports that Tregs suppress CD8 T-cell activation, we found that the (aTreg+rTreg):(Fr III) ratios were inversely correlated with the frequencies of CD8+ HLA-DR+ and CD8+ CD38+ cells(r=−0.399, p<0.001; r=−0.416, p<0.001; figure 4E,F). More importantly, the (aTreg+rTreg):(Fr III) ratios were significantly correlated with FEV1 predicted values (r=0.435, p<0.001; figure 4D), which indicated that the imbalance between suppressive subpopulations and the cytokine-secreting population was correlated with effector T-cell activation and severity of COPD.
Disturbed homeostasis among subpopulations of Tregs in BAL
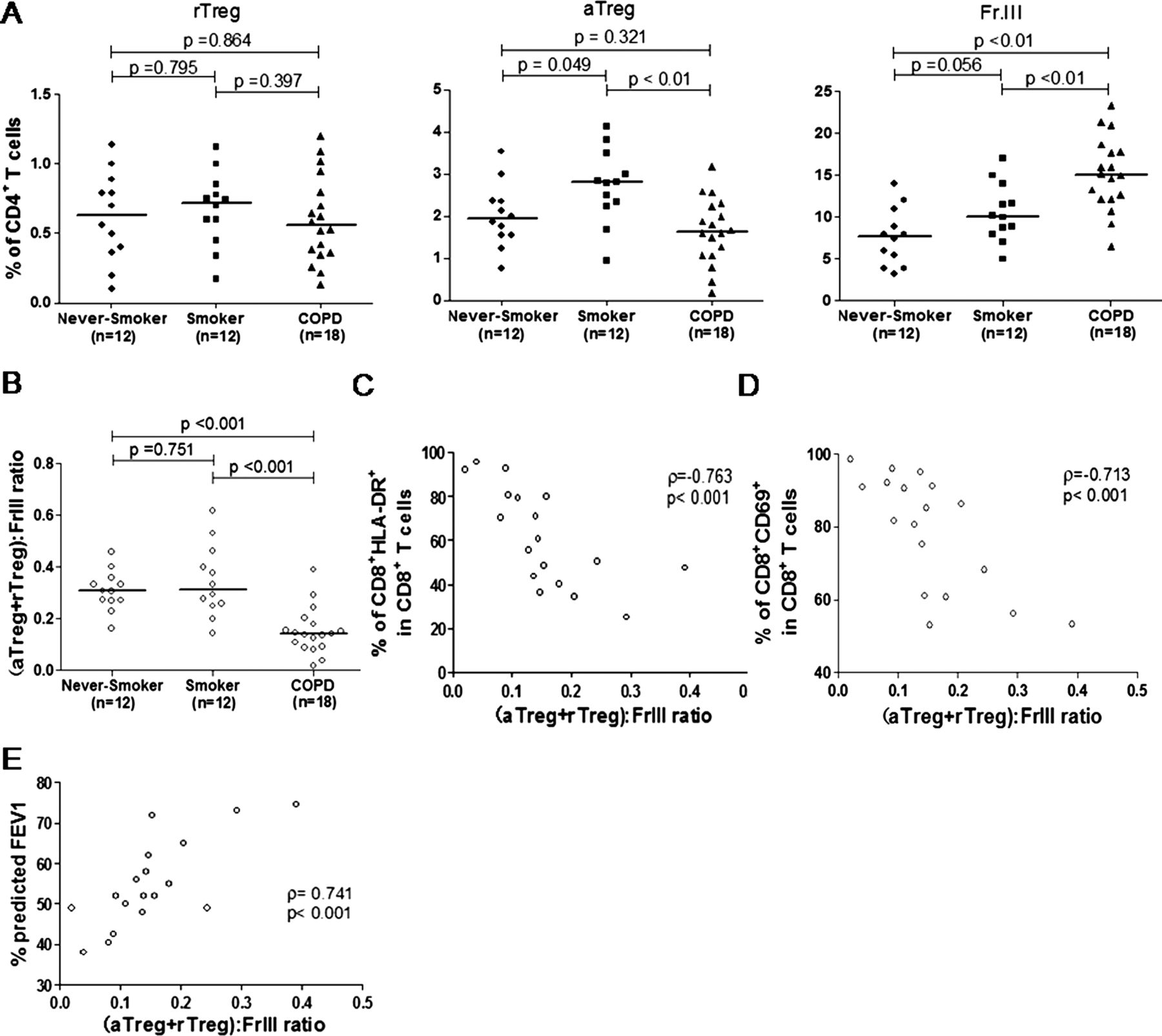
To clarify the homeostatic status of local Tregs, BAL samples were obtained from 12 never-smokers, 12 smokers and 18 patients with COPD who were undergoing clinically investigational bronchoscopy. In line with a previous study,18 we found that the majority of Tregs in BAL were CD45RA−. Accordingly, the percentage of rTregs in BAL was decreased markedly compared with that in peripheral blood and was identical across all three groups. However, aTregs and FrIII cells in BAL had a similar pattern of variation in peripheral blood, that is, aTregs decreased whereas Fr III cells increased in patients with COPD compared with never-smokers and smokers (figure 5A). Next, we calculated the (aTreg+rTreg):(Fr III) ratios in BAL to illustrate the local balance between the suppressive and pro-inflammatory subpopulations. Similar to the results from circulating Tregs, we found that (aTreg+rTreg):(Fr III) ratios were significantly lower in BAL of patients with COPD compared with smokers and never-smokers, suggesting that disturbed Treg homeostasis was also present in the lung (figure 5B). More importantly, compared with the blood (aTreg+rTreg):(Fr III) ratios, the BAL ratios showed a stronger correlation with activation status of effector T cells (CD8+ HLA-DR+: r=0.763, p<0.001; CD8+ CD69+ cells; r=0.713, p<0.001) and FEV1% predicted value (r=0.741, p<0.001; figure 5C–E).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The percentages of each Foxp3+ subset among CD4 T cells (A) and (aTreg+rTreg):(Fr III) ratios (B) in the bronchoalveolar lavage of never-smokers, smokers and patients with chronic obstructive pulmonary disease (COPD). Horizontal lines indicate median values. A p value<0.017 was considered significant. The (aTreg+rTreg):(Fr III) ratios correlated with CD8 T-lymphocyte activation status (C and D) and forced expiratory volume in 1 s (FEV1) predicted values (E) in patients with COPD. A p value<0.05 was considered statistically significant. Fr I, CD25++ CD45RA+ resting T-regulatory cells (rTreg); Fr II, CD25+++ CD45RA− activated T-regulatory cells (aTreg); Fr III, CD25++ CD45RA− cytokine-secreting cells.
Discussion
CD4+ Foxp3+ Tregs have been recognised as a negative regulatory population during immune responses.10 ,11 ,12 However, phenotypic and function heterogeneity of human CD4+ Foxp3+ Tregs is a major obstacle for understanding their clinical relevance. Dissecting CD4+ Foxp3+ cells into subsets, especially defining the cytokine-producing population (Fr III), is one of the striking findings of the studies on human CD4+ Foxp3+ Treg heterogeneity.21 ,24 ,25 Previous studies which identified Tregs as a whole did not find any significant difference in frequencies of circulating Tregs in patients with COPD.17 ,18 Here, based on this new definition of Treg subsets, we revealed significantly decreased proportions of rTregs and aTregs and an increased proportion of the cytokine-secreting non-Treg subpopulation in patients with COPD compared with smokers with normal lung function. More importantly, we linked the imbalance between these subsets with activation of CD8 T cells and the severity of COPD.
Despite a debate about their immune suppressive capacity, the inflammatory role of Fr III cells is well accepted due to their ability to produce IL-17 and IFN-γ. In smokers, we observed increased frequencies of three subpopulations, which suggested coexistence of anti-inflammatory and inflammatory responses. Strikingly, in comparison to smokers, patients with COPD exhibited increased proportions of Fr III cells and decreased proportions of rTregs and aTregs. This might reflect progression of inflammation and exhaustion of anti-inflammatory responses during disease progression.
Moreover, we further described an imbalance between anti-inflammatory and pro-inflammatory subsets by measuring the (aTreg+rTreg):(Fr III) ratio. Despite the elevation of three tested subsets in smokers with normal lung function, the (aTreg+rTreg):(Fr III) ratio remained stable as in never-smokers, which indicated that immune response homeostasis remained intact, although inflammatory and anti-inflammatory mechanisms were activated. However, in patients with COPD, as indicated by a significantly decreased (aTreg+rTreg):(Fr III) ratio, the balance between subpopulations tilted in favour of the inflammatory response, which might have facilitated inflammation-related lung injury. More importantly, in line with a decreased (aTreg+rTreg):(Fr III) ratio, we found a positive correlation between the circulating (aTreg+rTreg):(Fr III) ratio and FEV1%, which further linked the immune imbalance with airflow limitation in patients with COPD. The stronger correlation between the BAL (aTreg+rTreg):(Fr III) ratio and FEV1% provided more evidence to support this link. From these findings we hypothesise that a decrease in the immunosuppressive Treg populations, together with enhanced pro-inflammatory responses, induced by long-term exposure to inhalation of particles or gases, for example, tobacco smoking,18 lead to persistent airway inflammation dominated by CD8 T cells. This persistent airway inflammation is believed to be involved in the progressive loss of lung function characteristic of COPD.
It has been shown that CD8 T cells predominate over CD4 T cells in the airways and lung parenchyma of patients with COPD.1 ,5 COPD severity is correlated significantly with the frequency of activated CD8 T cells in peripheral blood.33 Moreover, previous studies have demonstrated opposing roles for Th17 cells and Tregs in CD8 cell activation: Treg cells suppress the proliferation of activated CD8 T cells via a variety of mechanisms,10–15 whereas IL-17-secreting cells promote CD8 cell activation by inducing expression of IL-6, IL-8 and intracellular adhesion molecule 1.34 ,35 It has been shown that BAL samples from patients with COPD also exhibit a higher CD8 cell:Treg ratio than smokers.17 In this study, we revealed a negative correlation between the circulating (aTreg+rTreg):(Fr III) ratio and frequency of activated CD8 T cells, which indicated that such enhanced activation of CD8 T cells might have resulted from immune imbalance in patients with COPD. Moreover, circulating CD8 T cells in patients with COPD exhibited elevated frequencies of cells expressing CD38 and HLA-DR, two general markers for T-cell activation, but not a transient marker CD69, which further indicated that activation of CD8 T cells in patients with COPD was a result of chronic inflammation rather than acute activation. Interestingly, pulmonary CD8 T cells in patients with COPD exhibited elevated expression of CD69 and HLA-DR, suggesting ongoing activation of T cells at the site of inflammation.
In summary, this study provides further evidence for the role of adaptive immunity in COPD pathogenesis, and demonstrates that the balance between the subpopulations of previously described CD4+ Foxp3+ Tregs may contribute to the progression of inflammation in the lung. Of note, the correlations between the (aTreg+rTreg):Fr III ratio and activated CD8 T cells or FEV1% existed in BAL, and to a lesser extent, in peripheral blood, indicating a closer correlation between loss of lung function and local immune activation. Furthermore, our data extend local immune activation to systemic immune activation, which might facilitate our understanding of the underlying mechanisms for local and systemic inflammation in COPD. Future therapeutic strategies to control inflammatory responses in COPD may be directed at modulation of different subpopulations of CD4+ Foxp3+ cells to restore immune homeostasis.
Acknowledgments
The authors thank Husheng Zhang, Dongning Chen, Ran Li, Yang Wang, Li Li and Haiyan Sheng (Beijing Tongren Hospital) for their support with screening participating subjects throughout the study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online appendix
- Data supplement 2 - Online figures
Footnotes
-
Contributors JH: contributed to recruiting the patients, performing all data collection, collecting and processing samples, and writing the manuscript. YS: contributed as primary investigator and was responsible for designing the study and writing the manuscript. YH and JH: performed laboratory-based assays. JZ, XL, PB and XZ: recruited the patients and collected clinical data. HZ: contributed as lead investigator and was responsible for designing the study, analysing the data and writing the manuscript. All authors read and approved the final manuscript.
-
Funding This work was supported by the Beijing Natural Science Foundation (No. 7092022), National Natural Science Foundation of China (No. 81170039) and 215 program (2009-2-13).
-
Competing interests None.
-
Ethics approval Ethics Committees, Beijing Tongren Hospital, Capital Medical University.
-
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Chest clinic