Article Text

Abstract
Background Respiratory syncytial virus (RSV) infection of airway epithelial cells (AECs) is an important initial event in RSV bronchiolitis. AEC immunological responses are thought to be critical in driving the subsequent inflammation in the airway. This study examined viral replication, cytotoxicity and cytokine production in cultures of primary AECs from children compared with responses to RSV infection in an immortalised epithelial cell line and to those from infants with RSV bronchiolitis.
Methods RSV replication, proinflammatory cytokine responses and cytotoxicity in RSV-infected primary AEC cultures derived from bronchial brushings from the lungs of children were compared with those seen in BEAS-2B cultures, as well as AECs and bronchoalveolar lavage fluid collected from children with and without RSV bronchiolitis.
Results Viral replication, cytotoxicity and inflammatory cytokine production were greater in primary AEC cultures than in BEAS-2B cells. Different response patterns were observed, with RSV infection of primary AEC cultures causing distinct peaks of viral replication and matched cytotoxic responses. Some primary AEC culture immunological responses, such as interleukin 8, were similar in magnitude to those seen in clinical samples from the lungs of children with RSV bronchiolitis. Although variable amounts of RSV were detected by PCR in freshly isolated primary AECs, RSV was not detected by immunocytochemistry.
Conclusion This is one of the first studies to examine comprehensively the responses to RSV infection in primary AEC cultures from children and shows marked differences from those of a commercially available immortalised human cell line but reassuring similarities to results found in vivo. This suggests that future work investigating responses of AECs to RSV infection should use primary AEC cultures.
- Bronchiolitis
- airway epithelium
- cell culture
- RSV infection
- broncho-alveolar lavage
- respiratory infection
- viral infection
- cytokine biology
- paediatric lung disaese
- paediatric physician
Statistics from Altmetric.com
- Bronchiolitis
- airway epithelium
- cell culture
- RSV infection
- broncho-alveolar lavage
- respiratory infection
- viral infection
- cytokine biology
- paediatric lung disaese
- paediatric physician
Key messages
What is the key question?
Do responses to RSV infection in commercially available immortalised epithelial cell line cultures adequately reflect those found in primary paediatric airway epithelial cell (pAEC) cultures, or those found in infants with RSV bronchiolitis?
What is the bottom line?
Responses to RSV infection found in primary paediatric airway epithelial cell cultures differ significantly from those found in immortalised cultures but show some similarities to those found in vivo, suggesting that future investigators examining AEC responses to RSV infection should consider using primary paediatric cell cultures.
Why read on?
This is one of the first studies to comprehensively examine the responses to RSV infection in pAEC cultures from children.
Introduction
Respiratory syncytial virus (RSV) is a common cause of acute respiratory infection (ARI) in both adults and children.1 2 In adults its disease burden is primarily in elderly individuals where it is now recognised as being similar in magnitude to that of seasonal influenza.3 4 In children RSV causes bronchiolitis, a leading cause of morbidity and mortality worldwide.5 In Europe, RSV is associated with approximately 40% of hospital admissions for acute respiratory infection in children <2 years of age, with the youngest infants experiencing the most severe disease.6 Supportive therapy (oxygen and/or fluids) has been the mainstay of treatment for the majority of children with bronchiolitis for the last 50 years.7 Prevention of RSV disease is now possible, but this is only available for infants at high risk of severe disease.8 9 Attempts to develop a vaccine to prevent RSV infection have so far been unsuccessful.
The lack of effective treatments or vaccines makes understanding the pathogenesis of RSV disease all the more important. In the airway it is thought that infection of airway epithelial cells (AECs) by RSV initiates the inflammatory responses which characterise bronchiolitis through the release of early inflammatory mediators such as tumour necrosis factor α (TNFα), eotaxin and interleukin 8 (IL-8).10 11 Although it has not been possible to study the specific contribution of AECs to the immune response in infants with RSV bronchiolitis in vivo, murine studies,12–14 air-liquid interface cultures15–18 and immortalised cell lines19 20 have provided useful information, although each of these models has drawbacks. Air-liquid interface cultures, for example, are not easy to develop and generally require the use of post-mortem material, while murine studies and immortalised cell lines may not accurately reflect responses to infection in humans.
In this study we have used a cell culture model derived from primary AECs from children and assessed its utility as a model of RSV infection. We have compared RSV replication, cytokine responses and cytotoxicity with those seen in an immortalised bronchial epithelial cell line commonly used to study viral-epithelial interactions. We have also compared responses seen in this model with those found in freshly isolated primary AECs and bronchoalveolar lavage (BAL) fluid from children with and without RSV bronchiolitis.
Methods
Patients
Bronchial brushings and BAL fluid were obtained from healthy children undergoing elective surgery for non-respiratory conditions. Samples were also obtained from clinically stable infants ventilated with RSV bronchiolitis, born at term with no underlying medical problems.
Sample collection
Non-bronchoscopic bronchial brushing was performed using an unsheathed nylon cytology brush as described previously.21 Briefly, the brush was inserted directly through the endotracheal tube until resistance was felt and then rubbed gently against the epithelial surface. Once the brush had been withdrawn, we cut the tip cut-off into 5 ml of collection media and transported the sample to the laboratory. Following centrifugation of brush and media (1300 rpm; 350 g), the brush and media were discarded and a portion of the cell pellet was resuspended in phosphate-buffered saline (PBS) for cytospin preparation; part of the remaining cell pellet was snap frozen for protein analysis and the remainder was used for cell culture.
Non-bronchoscopic BAL fluid was collected from children with and without RSV bronchiolitis, as described previously.22 A suction catheter was passed down the endotracheal tube. Two 1 ml/kg aliquots of sterile normal saline were instilled separately down the suction catheter. BAL fluid was recovered with constant suction pressure into a mucus trap and then processed according to ERS guidelines.
AEC culture
Primary AECs were cultured from healthy children until the second passage as described previously.17 Both primary AECs and BEAS-2B cells (European Cell Culture Collection) were grown in supplemented bronchial epithelial growth medium Lonza (Wokingham, UK) on flasks coated with 0.01 mg/ml fibronectin Sigma-Aldrich (Dorset, UK), 0.03 mg/ml collagen BD Biosciences (Oxford, UK) and 0.01 mg/ml bovine serum albumin (Sigma-Aldrich).
Virus
We cultivated and harvested RSV A2 (gift from Professor S L Johnson, Imperial College, London) as described previously.23 Briefly, virus was added to Hep-2 monolayers grown in flasks with DMEM supplemented with 10% heat-inactivated fetal calf serum. Cells were incubated with the virus until the cytopathic effect reached approximately 50% (48 h). The monolayer was then harvested, centrifuged at 500 g and lysed on ice. Aliquots were snap frozen and stored at −70°C.
The RSV titre was determined using a modified plaque-forming unit assay. Briefly, the virus was serially diluted and added to Hep-2 cell monolayers grown in 96-well plates (total volume 50 μl). The cells were incubated for 2 h before being fixed in methanol and stained with an anti-RSV biotinylated antibody (goat anti-RSV, AbD Serotec (Oxford, UK)) labelled using an extravidin peroxidise colour development substrate (Sigma Fast-Red, Sigma-Aldrich). Plaques were visualised using light microscopy and counted. Plaque forming units (PFU) were calculated as: number of plaques/ml × dilution factor × 20 (for 50 μl culture volume).
The RSV-specific nature of the responses observed was confirmed by inhibiting virus-induced IL-6 responses in RSV-infected BEAS-2B cultures treated with increasing concentrations of palivizumab (Abbott Laboratories, Maidenhead, UK) (a commercially available monoclonal antibody against RSV F-protein). Dilutions of 4, 400 and 1600 μg/ml of palivizumab were prepared and added to 0.25 MOI RSV A2. This mixture was then added to the culture and the cultures were harvested at 24 h post-inoculation (see online supplement).
Immunocytochemistry
Epithelial cells from RSV-infected and non-infected cultures were spun onto glass microscope slides, fixed in 10% neutral buffered formalin, washed in PBS and stored at −20°C prior to analysis. After rehydrating in Tris buffered saline, cytospins were blocked with 1% goat serum in Tris buffered saline and then stained with the epithelial marker cytokeratin-19 using a mouse anti-human monoclonal antibody (BD Biosciences) tagged with a Texas red goat anti-mouse secondary antibody (Sigma-Aldrich).21 We also stained cells with a directly conjugated (FITC) mouse monoclonal antibody for RSV nucleoprotein Abcam (Cambridge, UK). Finally, 4′,6-diamidino-2-phenylindole (DAPI) was used to stain cell nuclei. Cytospins were mounted using fluorescence mounting media Dako (Ely, UK) before being visualised by confocal microscopy Leica (Milton Keynes, UK).
Cytospins of freshly isolated primary AECs and BAL fluid from healthy and RSV-infected patients were prepared and stained in the same way.
Viral replication
Primary AECs from healthy children and BEAS-2B cells were cultured to 80% confluence and then infected with varying titres (0.0025, 0.025, 0.25 and 2.5 MOI) of RSV. Viral replication was measured using a commercially available TaqMan real-time PCR kit that quantified PFU from a known standard curve measuring RSV N-gene expression to which interpolation of sample values could then be made Primer Design (Southampton, UK). This was completed for all cultures at 3, 6, 12, 24 and 48 h after infection. Total RNA was extracted using RNeasy kit Qiagen (West Sussex, UK) and 2 μg used as a template for cDNA synthesis using random primers and M-MLV reverse transcriptase Promega (Southampton, UK). We determined RSV N-gene expression by TaqMan real-time PCR and normalised to expression of the housekeeping gene cyclophilin A (PPIA) Applied Biosystems (Warrington, UK).24
Viral replication in freshly isolated AECs and BAL fluid from infants with bronchiolitis was similarly assessed using RSV N-gene expression and normalised to, respectively, PPIA and another housekeeping gene, L32 (Applied Biosystems).
Cytotoxicity
Lactate dehydrogenase (LDH) release was used as a measure of cytotoxicity in culture supernatants and BAL fluid. Supernatant from BEAS-2B cells treated with 0.5% Triton X for 24 h was used as a ‘high’ LDH reference control, according to the manufacturer's instructions Roche (West Sussex, UK). LDH activity in all samples was then calculated as a percentage of the total cellular LDH released in this ‘high’ control sample as previously described.25 26 Levels of <10% were considered to be within the normal range, while levels of >100% indicated cytotoxicity greater than that seen in the ‘high’ LDH reference control.
Cytokine production
We measured secreted IL-6 and IL-8 in cell culture supernatants and BAL fluid using commercially available ELISA kits R&D Systems (Abingdon, UK) according to the manufacturer's instructions (Quantipro, Sigma-Aldrich).
Statistical analysis
Data from RSV-infected and healthy patient samples were expressed as medians (IQR) or means (SD) depending on their distribution, with statistical differences between groups assessed using the Mann–Whitney U test or Student t test, respectively.
All in vitro experiments were completed in duplicate with a minimum of three biological replicates. Duplicate results were averaged to give an experimental value that was averaged across replicates. The final mean replicate values with SEM are shown unless otherwise stated. Infected and non-infected cultures were tested for differences using the Student t test with significance taken at p<0.05.
Results
Clinical characteristics
Bronchial brushings and BAL fluid were collected from 20 healthy non-atopic children (nine boys) of median age 8 years (range 2–15) undergoing elective surgery for non-respiratory conditions (see table in online supplement). Non-atopic status was based on a normal serum IgE and a negative radioallergosorbent test (RAST) to common aeroallergens. The absence of respiratory symptoms was confirmed by negative responses on the ISAAC and ATS respiratory questionnaires.
Samples were also collected from 13 clinically stable infants (six females) of median age 4 weeks (range 2–37) ventilated with RSV bronchiolitis, born at term with no underlying medical problems. RSV status was ascertained by enzyme-linked fluorescence Biomerieux (Basingstoke, UK) of nasopharyngeal aspirate. The parents of seven infants gave consent for both bronchial brushing and BAL while the parents of six infants gave consent for only BAL to be performed. We collected samples within 24 h of admission, just prior to routine bronchial toilet, and sample collection was well tolerated by all children. We were unable to perform all analyses on all samples due to variations in sample size. The mean (SD) bronchial brushing cell yield from healthy children and those with RSV was 3.4 (1.2)×106 and 2.6 (1.1)×106 cells, respectively. The mean (SD) BAL cell yield from healthy children and those with RSV was 2.1 (0.2)×106 cells and 5.3 (3.0)×106 cells, respectively (see table in online supplement).
RSV infection in vitro
Viral replication
Primary AEC and BEAS-2B cell cultures infected with RSV were assessed for the presence of RSV and differences in RSV replication. Similar staining patterns for RSV protein were observed in cytospins of infected primary AECs and BEAS-2B cells (data not shown). The presence of replicating RSV was quantified with a commercially available kit using real-time PCR analysis of RSV N-gene expression. These values were interpolated to a standard curve for an indication of viral PFUs. RSV replication was much greater in primary AECs than in BEAS-2B cultures, particularly for RSV titres of >0.025 MOI (figure 1A). In primary AEC cultures a biphasic pattern in viral replication was seen with the first peak at 6 h (1–2×105 PFU), a trough occurring between 12 and 24 h and a second rise occurring at 48 h (4–5×105 PFU) following infection. In BEAS-2B cultures the presence of RSV peaked once at 12 h but at levels 2–3 log-fold less than in the primary cultures (2–8×102 PFU).

(A) Respiratory syncytial virus (RSV) replication in (i) primary airway epithelial cell (AEC) and (ii) BEAS-2B cultures over time. RSV N-gene expression was measured in infected and non-infected airway epithelial cultures by real-time PCR and normalised against L32 gene expression. These values were quantified as plaque forming units (PFU) by interpolation to a commercially supplied standard curve (n=3). Different phases of infection are seen in the primary cell culture model but not the immortalised cell line. (B) RSV-induced cytotoxicity in (i) primary AEC and (ii) BEAS-2B cultures over time. Lactate dehydrogenase (LDH) activity was measured in supernatants collected from RSV-infected and non-infected primary AECs and BEAS-2B cells using a commercially available assay. Absorbance values were compared with a known standard curve and sample values calculated by interpolation. These are presented as a percentage of LDH activity measured in BEAS-2B cells treated for 24 h with 0.5% Triton X used as a reference ‘high’ control (n=3). Different phases of infection are again seen in the primary cell culture model.
Cytotoxicity
LDH was measured in supernatants collected from RSV-infected primary AEC and BEAS-2B cell cultures to assess cytotoxicity caused by infection. The pattern and magnitude of the cytotoxic response to RSV infection differed in primary AEC and BEAS-2B cultures (figure 1B). In primary AEC cultures, biphasic responses were seen with peaks over 100% at 6 and 24 h following 2.5 MOI RSV infection. Similar response patterns were also seen at lower MOIs.
In BEAS-2B cultures the cytotoxic response to RSV infection remained below 10% at most time points and MOIs, apart from 6 and 48 h at 2.5 MOI RSV where the maximum cytotoxic response was 40%.
Cytokine responses
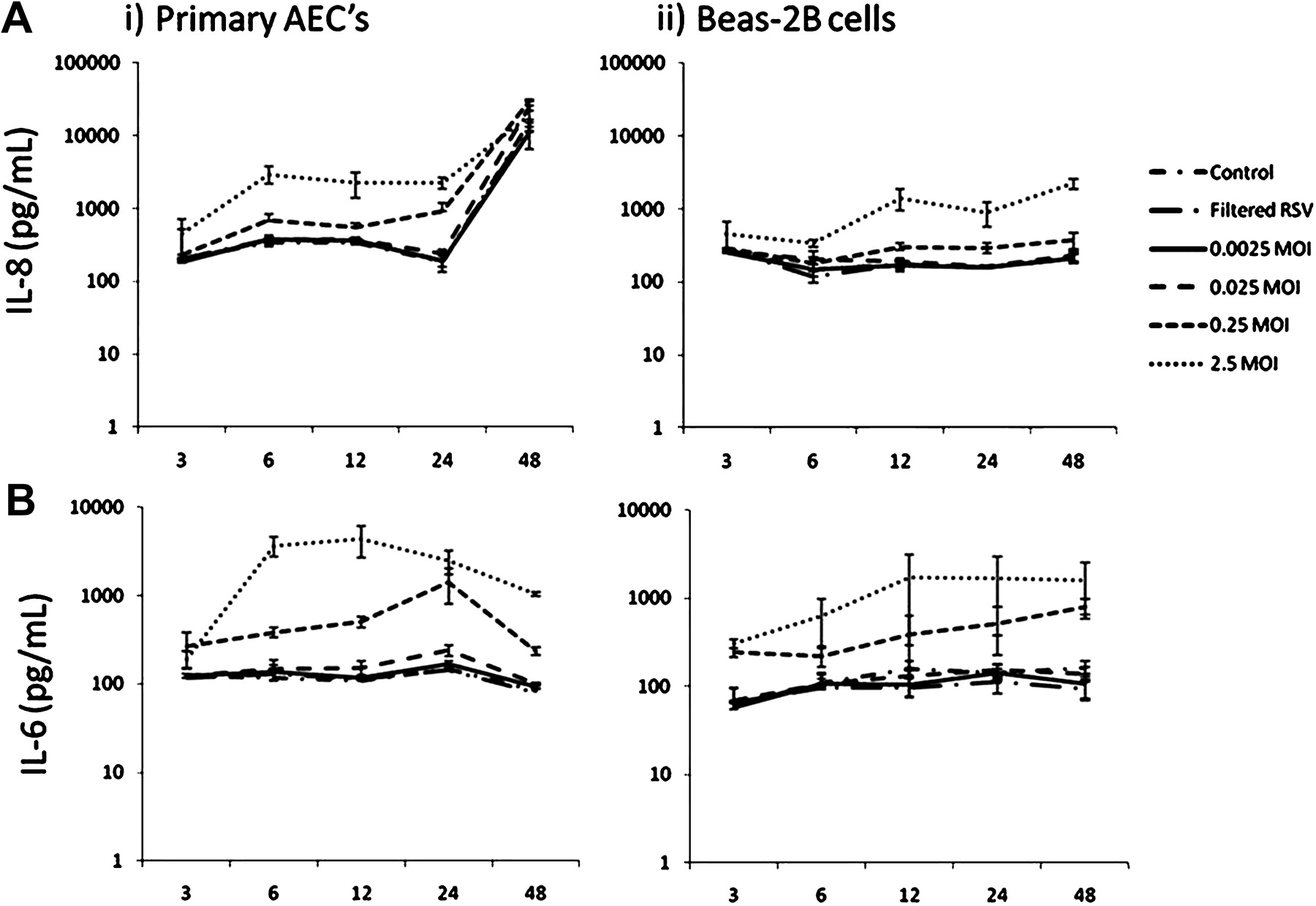
Secreted IL-8 was raised after 6 h in supernatants from RSV-infected primary AEC cultures and then rose again precipitously (in both RSV-infected and non-infected cultures) at 48 h following infection (figure 2). IL-8 was significantly greater in primary AEC cultures infected with 0.25 and 2.5 MOI RSV at 6, 12 and 24 h following infection than in controls or lower MOIs of RSV.
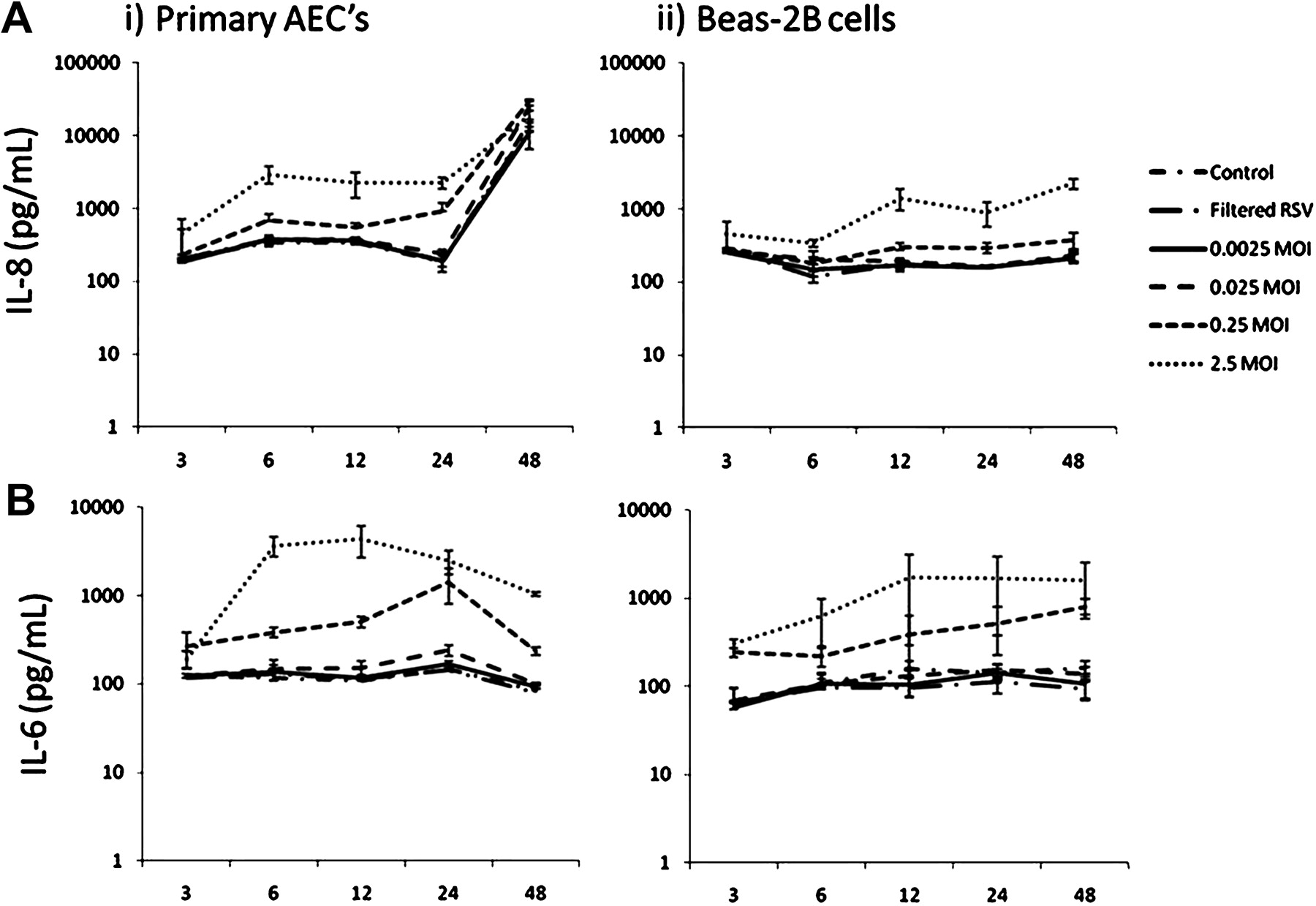
(A) Respiratory syncytial virus (RSV)-induced interleukin 8 (IL-8) production in (i) primary airway epithelial cell (AEC) and (ii) BEAS-2B cultures over time. Primary AECs secreted larger amounts of IL-8 than BEAS-2B cells in response to RSV infection at titres >0.25 MOI for all time points. A significantly greater IL-8 response was measured at 48 h in primary AECs for all treatments including controls; this was attributed to overconfluence. (B) RSV-induced interleukin 6 (IL-6) production in (i) primary AEC and (ii) BEAS-2B cultures over time. Primary AECs secreted larger amounts of IL-6 than BEAS-2B cells in response to RSV infection at titres >0.25 MOI for incubation times >6 h (n=3).
IL-8 concentrations in BEAS-2B cultures were less than those in primary AEC cultures and no steep rise was seen at 48 h following infection. IL-8 was greater in BEAS-2B cultures infected with 0.25 and 2.5 MOI RSV at 12, 24 and 48 h than in controls or lower MOIs of RSV.
IL-6 concentrations in culture supernatants were higher in primary AEC than BEAS-2B cultures at higher MOIs of RSV (figure 2). IL-6 responses were greatest 6–24 h following infection with 0.25 and 2.5 MOI RSV. IL-6 concentrations in primary AECs decreased in all groups at 48 h. Basal IL-8 and IL-6 production of >500 pg/ml was observed in both primary AEC and BEAS-2B cultures in all groups at all time points and MOIs.
Patient samples
Viral replication
The presence of RSV in freshly isolated AECs collected from RSV-infected and healthy children were assessed for comparison with the cell culture models. Interestingly, RSV protein was not detected in freshly isolated AECs (cytokeratin-19 positive) from infants with RSV bronchiolitis (figure 3). In contrast, BAL fluid from infants with RSV bronchiolitis contained cells, often in clumps or as syncytia, which stained for both cytokeratin-19 and RSV protein.
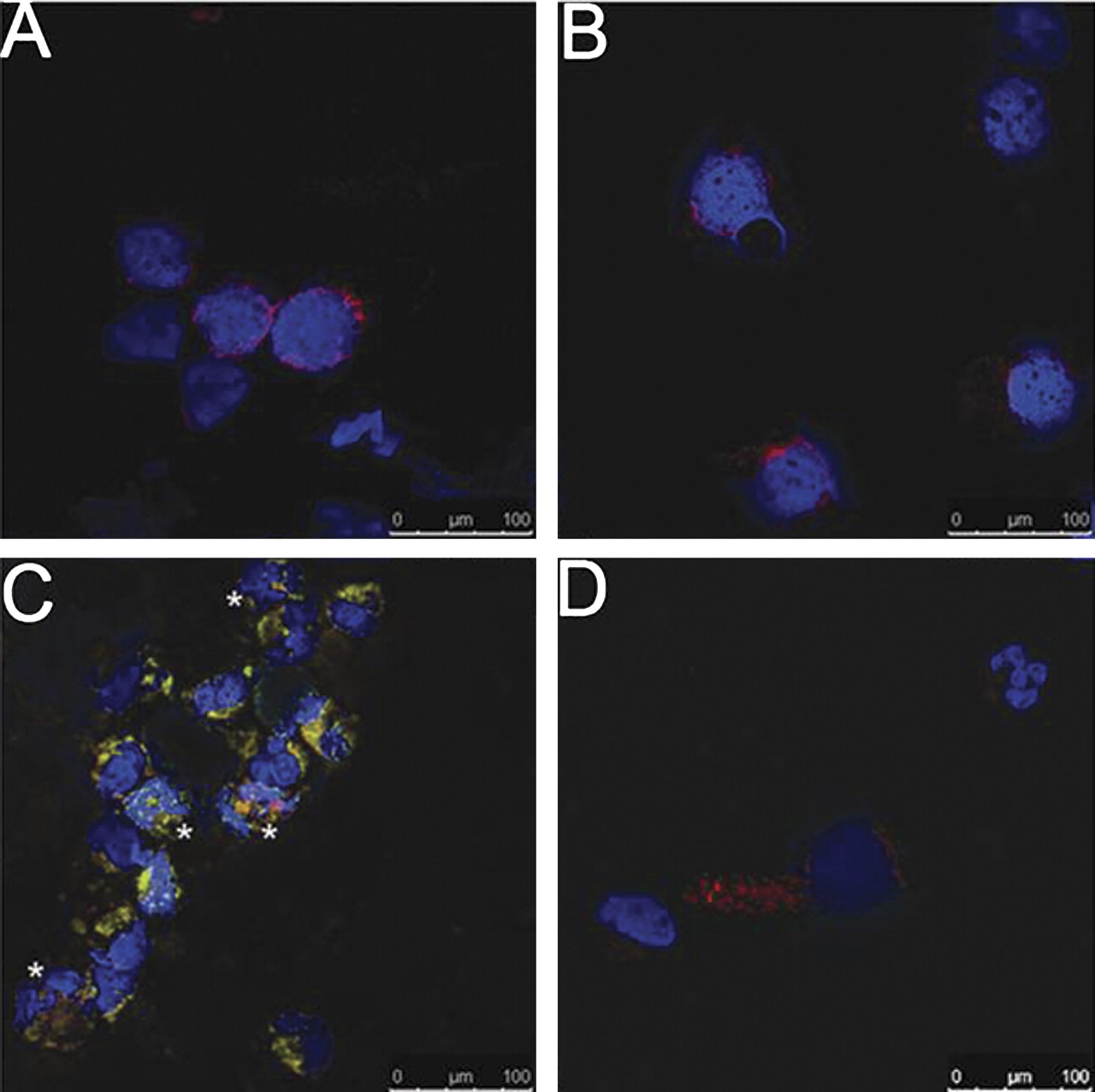
Confocal microscopy of respiratory syncytial virus (RSV) infection in (A, B) freshly isolated airway epithelial cells (AECs) and (C, D) bronchoalveolar lavage fluid from children with (A, C) and without (B, D) bronchiolitis. RSV protein (FITC) was not observed in AECs (Texas red) collected by bronchial brushing from children with (n=5) or without (n=11) RSV bronchiolitis (A, B respectively). RSV protein was observed in AECs in the predominantly neutrophilic bronchoalveolar lavage fluid collected from RSV-infected children (C; asterisks adjacent to infected AECs) but not from healthy children (D). Cell nuclei shown in blue (DAPI) (staining representative of n=3 for each sample type).
The amount of replicating RSV present was quantified (PFU) using real-time PCR analysis of RSV N-gene expression. Figure 4 shows that the presence of RSV in samples of freshly isolated AECs and BAL fluid from infants with RSV bronchiolitis varied considerably (AEC: median 1.28 (range 0.03–106.74)×103 PFU; BAL: median 5.362 (range 0.003–341.68)×103 PFU). In one infant with RSV bronchiolitis, RSV-B (rather than RSV-A) N-gene was detected in both freshly isolated AECs and BAL fluid. RSV was not detected in healthy non-atopic control patients.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Viral replication, cytotoxic and inflammatory responses in freshly isolated airway epithelial cells (AECs) and bronchoalveolar lavage (BAL) fluid from healthy children and infants with respiratory syncytial virus (RSV) bronchiolitis. (A) RSV N-gene expression in AECs (n=5) and BAL fluid (n=8) from infants with RSV bronchiolitis (black bars represent median values). RSV N-gene expression was not detected in healthy control patients (data not shown). (B) Lactate dehydrogenase (LDH) activity measured in BAL fluid collected from healthy children and infants with RSV bronchiolitis (healthy=5; RSV=11). (C) Interleukin 8 (IL-8) and (D) interleukin 6 (IL-6) protein in AECs and BAL fluid from healthy and RSV-infected patients. *p=0.019.
Cytotoxicity and cytokine analysis
BAL fluid collected from RSV-infected and healthy patients was assessed for indicators of cell death (LDH) and inflammation (IL-6 and IL-8). Figure 4 shows LDH in BAL fluid from infants with and without RSV bronchiolitis. While five infants had significant LDH, no LDH was detected in six infants (range 0–293%). LDH was not detected in BAL fluid from healthy non-atopic children.
IL-6 and IL-8 concentrations in cell pellets were similar in AECs from children with and without RSV bronchiolitis (figure 4). IL-8 concentrations in BAL fluid from infants with RSV bronchiolitis were greater than those in BAL fluid from healthy non-atopic children (p=0.019). BAL fluid IL-8 but not IL-6 levels were similar to those seen in RSV-infected primary AEC cultures (>0.025 MOI).
Discussion
This is the first study to compare RSV infection in primary monolayer cultures derived from epithelial cells from the lungs of children with an immortalised cell line commonly used to investigate pulmonary responses to viral infection. Compared with primary AEC cultures, BEAS-2B cells were relatively resistant to RSV infection and demonstrated much lower levels of viral replication, cytotoxicity and inflammatory cytokine production. Different patterns of infection were also observed, with RSV causing two distinct peaks of viral replication with matched cytotoxic responses in primary AEC cultures. Although the magnitude of some cytokine responses in primary AEC cultures were similar to those in samples from the lungs of children with RSV bronchiolitis, no such similarity in viral replicative and cytotoxic responses was seen.
RSV infection was more efficient in our primary AEC model, with RSV replication and cytotoxicity at higher RSV MOIs being 100 and 15 times greater, respectively, than in BEAS-2B cells. Similar patterns were also seen with IL-8 and IL-6 responses. While this has not been demonstrated before, Donnelly et al showed that granulocyte-macrophage colony stimulating factor responses to cytomix (IL-1β, TNFα and interferon-γ) stimulation were greater in freshly isolated adult AECs than in passaged adult AEC cultures, and that both were greater than those seen in A549 and BEAS-2B cells.27 Possible reasons for this difference between primary and immortalised cultures might include variations in cell surface receptor expression for RSV.28 Equally, RSV infection might induce different antiviral responses in BEAS-2B cells compared with primary AEC cultures because of changes undergone during transfomation/immortalisation.
Laham et al have recently proposed nasopharyngeal LDH as a possible predictor of disease severity, with high LDH levels being associated with decreased risk of hospitalisation in infants with RSV bronchiolitis.29 The source of LDH was speculated upon in this study, but we clearly demonstrate that RSV-infected epithelial cell cultures are capable of producing large amounts of LDH. Infection with low RSV MOIs resulted in LDH activity being higher at 24 h than 6 h, while LDH activity with 2.5 MOI RSV was greater at 6 h than 24 h. It is likely that these subtle differences reflect the number of cells available for progeny virus to infect following the first wave of infection. The second cytotoxic peak appearing to precede the ‘second’ viral replication peak may reflect our sampling schedule which might have missed another replication peak between 12 and 24 h.
This is the first study to examine AEC responses in infants with RSV bronchiolitis. Our findings appear to contradict each other, with RSV being detected in brushed AECs by PCR but not on immunocytochemistry. While this may be due to lower sensitivity of immunocytochemistry, it is also possible that the airway epithelium sampled may not have been infected with RSV, and that any RSV detected by PCR may have been free virus picked up within the airway. This inability to demonstrate overt RSV infection in AECs may also reflect the limitations of our sampling technique. In our experience from collecting brushings under bronchoscopic guidance in older children, we normally sample from fourth to sixth generation bronchi. However, in an infant weighing 4–5 kg, the cytology brushes used would probably have only reached second or third generation bronchi, some distance from the bronchioles. It is possible that these airways were either infected earlier in the course of the illness and had recovered or were not infected at all during the illness.
At 48 h, IL-8 concentrations were 100–1000 times higher than baseline levels in primary AEC cultures (but not in BEAS-2B cultures), whether infected with RSV or not. We believe this is for two separate reasons. In infected cultures, particularly at higher MOIs, primary AEC IL-8 production has either peaked or is peaking as all cells become infected. In uninfected cultures, elevated IL-8 production may be due to overconfluence of cells with resultant cell death as described previously.30 Some studies using primary AECs have even removed 48 h time points because of this phenomenon.31 For these reasons we believe that IL-8 is not the best indicator of RSV infection in confluent cultures incubated for more than 48 h.
In primary AEC cultures, at multiple time points and MOIs, we have characterised RSV replication and subsequent cytotoxic and cytokine responses, thereby determining optimal conditions (0.25 MOI RSV at 24 h) for future experiments. IL-8 responses in our model, in contrast to infected BEAS-2B cells, were also of the same order of magnitude as those seen in BAL fluid from children with RSV bronchiolitis (acknowledging the limitations of this method for sampling airway secretions). However, these primary cultures are based on APCs from young children and not infants aged <6 months in whom most of the severe RSV disease is seen. An examination of these responses in cultured epithelial cells from infants prior to any RSV infection could potentially clarify whether these responses differ with age and affect disease pathogenesis, but would be a difficult study to conduct. Recent studies have suggested that RSV A2 induces greater cytopathogenicity than wild-type RSV in culture.18 Our study provides another model in which to investigate these mechanistic differences.
In conclusion, we have shown that primary epithelial cells derived from bronchial brushings from the lungs of children differ markedly from a commonly used epithelial cell line in their responses to RSV infection. We would suggest that cell lines be used to determine mechanistic pathways, but that definitive experiments should be performed using primary cells to confirm any observed effects in a more relevant model.
References
Footnotes
Funding An Intermediate Clinical Fellowship for PSMcN was funded by the Wellcome Trust.
Competing interests None.
Patient consent Obtained.
Ethics approval The Liverpool paediatric research ethics committee approved the study and all samples were collected following written informed consent from parents.
Provenance and peer review Not commissioned; externally peer reviewed.