Article Text

Abstract
Background Emerging evidence shows that airway microbiota may modulate local immune responses, thereby contributing to the susceptibility and severity of acute respiratory infections (ARIs). However, there are little data on the longitudinal relationships between airway microbiota and susceptibility to ARIs in children.
Objective We aimed to investigate the association of early nasal microbiota and the subsequent risk of ARIs during the first years of life.
Methods In this prospective population-based birth-cohort study in Finland, we followed 839 healthy infants for ARIs from birth to age 24 months. Nasal microbiota was tested using 16S rRNA gene sequencing at age 2 months. We applied an unsupervised clustering approach to identify early nasal microbiota profiles, and examined the association of profiles with the rate of ARIs during age 2–24 months.
Results We identified five nasal microbiota profiles dominated by Moraxella, Streptococcus, Dolosigranulum, Staphylococcus and Corynebacteriaceae, respectively. Incidence rate of ARIs was highest in children with an early Moraxella-dominant profile and lowest in those with a Corynebacteriaceae-dominant profile (738 vs 552/100 children years; unadjusted incidence rate ratio (IRR), 1.34; 95% CI 1.16 to 1.54; p < 0.001). After adjusting for nine potential confounders, the Moraxella-dominant profile-ARI association persisted (adjusted IRR (aIRR), 1.19; 95% CI 1.04 to 1.37; p = 0.01). Similarly, the incidence rate of lower respiratory tract infections (a subset of all ARIs) was significantly higher in children with an early Moraxella-dominant profile (aIRR, 2.79; 95% CI 1.04 to 8.09; p = 0.04).
Conclusion Moraxella-dominant nasal microbiota profile in early infancy was associated with an increased rate of ARIs during the first 2 years of life.
- children
- infant
- lower respiratory tract infection
- moraxella
- nasal microbiota
- respiratory infections
- steps study
- wheezing
Statistics from Altmetric.com
- children
- infant
- lower respiratory tract infection
- moraxella
- nasal microbiota
- respiratory infections
- steps study
- wheezing
Key messages
What is the key question?
Are early infant airway microbiota profiles associated with susceptibility to acute respiratory infections (ARIs) in young children?
What is the bottom line?
In this prospective, population-based birth-cohort study, we identified five distinct nasal microbiota profiles in healthy 2-month-old infants, and found that the incidence rate of ARIs was highest in children with an early Moraxella-dominant nasal microbiota profile and lowest in those with a Corynebacteriaceae-dominant profile.
Why read on?
This large prospective cohort study shows that early infant airway microbiota profiles may play an important role in susceptibility to ARIs in children during the first years of life.
Introduction
Acute respiratory infection (ARI) is a major cause of morbidity in infants and young children with a mean annual rate of 5–7 infections per child.1–3 Clinical manifestations of ARI range from the self-limited common cold to severe (and even fatal) infections. Lower respiratory tract infections (LRTIs), including pneumonia and bronchiolitis, are the most frequent cause of hospitalisation in children, accounting for 50%–60% of hospitalisations before age 5 years.4 Some children are more susceptible to ARIs but known risk factors (such as age, household siblings, daycare and prematurity) do not completely explain the variability in the incidence and severity of ARIs.
The recent advent of high-throughput sequencing methods has identified diverse microbial communities in the airways. Emerging evidence shows that airway microbiota may modulate local immune responses5 6 thereby contributing to the susceptibility7–9 and severity of ARIs.10–12 Previous studies investigating the association between airway microbiota and susceptibility to ARIs in children have been small,7 9 11 conducted in selected subjects (eg, infants at high risk for atopy),11 have not investigated the rates of all types of ARIs,7 9 or the airway microbiota has been concurrent with ARI.8 10 12 13 Furthermore, studies have reported seemingly contradictory associations between airway microbiota and ARI incidence, specifically for Moraxella.7 9 11 14 It remains to be elucidated how airway microbiota in the early infancy relates to the subsequent risks of ARIs (and, more specifically, LRTIs) in healthy children. Better understanding of this relationship would inform the development of preventive measures for children at high risk for ARIs.
To address this knowledge gap, we conducted a prospective population-based birth-cohort study in healthy children to identify nasal microbiota profiles in the early infancy and to examine the association of these profiles with the rates of ARIs during age 2–24 months.
Methods
Study design and conduct
We analysed data from the prospective, population-based birth-cohort study—the Steps to the Healthy Development and Well-being of Children (STEPS Study)—where families of Finnish infants born in January 2008 through April 2010 were enrolled during pregnancy or soon after birth.15 As part of the STEPS study, 923 infants were enrolled in a follow-up for ARIs from birth to age 24 months.1 The details of the study design, setting, subjects, testing and analysis may be found in the online supplement. Briefly, parents were asked to record all respiratory and other symptoms and physician visits with associated diagnoses and treatments into a daily symptom diary. Families were instructed to visit the study clinic during ARIs if they felt that an evaluation by a physician was needed. At the study clinic, children were examined by a study physician and clinical findings were documented using a structured form. Data on outpatient and emergency department visits at hospitals and hospitalisations from birth to age 24 months were retrieved from medical records of the Hospital District of Southwest Finland. Patient demographics, family history, prenatal, perinatal, and postnatal history, and environmental information were collected from the National Birth Registry and by structured questionnaires. Infants were examined at the study clinic at age 2 months at healthy state and nasal swab specimens were collected by trained study personnel using flocked nylon swabs (Copan, Brescia, Italy). Any current respiratory symptoms were documented. Parents of participating children gave their written, informed consent. The study complies with the Declaration of Helsinki.
Supplemental material
Respiratory virus detection
The nasal swab samples were stored at −80°C after the collection. Swabs were suspended in phosphate-buffered saline and analysed by using real-time, quantitative PCR for rhinovirus, human enteroviruses and respiratory syncytial virus (RSV) as described earlier.16 17 All specimens collected during influenza seasons were analysed by reverse transcriptase PCR for influenza A and B viruses.18
16S rRNA gene sequencing of nasal airway microbiota
16S rRNA gene sequencing methods were adapted from the methods developed for the National Institutes of Health Human Microbiome Project.19 20 The details of the methods are described in the online supplemental methods. Briefly, bacterial genomic DNA was extracted with automated MagNA Pure 96 System (Roche Life Science). The 16S rDNA V4 region was amplified by PCR and sequenced on the MiSeq platform (Illumina, San Diego, California, USA) using the 2×250 bp paired-end protocol yielding pair-end reads that overlap almost completely. Sequencing reads were merged using USEARCH V.7.0.1090.21 Samples with suboptimal amounts of sequencing reads were resequenced to ensure that most bacterial taxa were encompassed in our analyses. 16S rRNA gene sequences were clustered into operational taxonomic units (OTUs) at a similarity cut-off value of 97% using the UPARSE algorithm.22 OTUs were determined by mapping the centroids to the SILVA database23 containing only the 16S V4 region to determine taxonomies. Rarefaction curves of bacterial OTUs were constructed using sequence data. We utilised multiple quality control measures, including the use of non-template controls in the microbial DNA extraction, 16S rRNA gene amplification and amplicon sequencing processes. The details of the quality control measures are described in the online supplemental methods. Analyses were conducted at the genus level.
Outcomes
The primary outcome was the incidence rate of all ARIs (including upper ARIs and LRTIs). The secondary outcomes were (1) the rate of LRTIs during age 2–24 months and (2) recurrent wheezing by age 24 months. An ARI was defined as the presence of rhinitis or cough (with or without fever or wheezing) documented in the symptom diary by the parents, or as any ARI diagnosed by a physician.1 The LRTIs (eg, bronchiolitis, pneumonia) were diagnosed by physicians. Recurrent wheezing was defined as ≥3 wheezing episodes during age 2–24 months as reported by the parents in the diary or diagnosed by a physician.
Statistical analysis
To identify nasal airway microbiota profiles at age 2 months, we performed unsupervised clustering by partitioning around medoids24 based on the Bray-Curtis distance at the genus level.25 The gap statistic method was used to select the number of profiles for the data (online supplementary figure E1).26 The patient characteristics and outcomes were compared between the derived nasal microbiota profiles, using the χ2, Fisher’s exact and Kruskal-Wallis rank sum test as appropriate. The relative abundance of bacterial genera was compared between the profiles by using analysis of variance adjusting for multiple comparisons with the use of the Benjamini-Hochberg false discovery rate method.27 Incidence rates of all ARIs and LRTIs were analysed by using unadjusted and multivariable-adjusted negative binomial regression models using subject-level data with natural logarithm of the individual completed follow-up time from age 2 month nasal sample to age 24 months as an offset. Risk of recurrent wheezing was analysed by modelling the time to event (ie, time from age 2 month nasal sample to the third wheezing episode) using Kaplan-Meier method, log-rank test and Cox proportional hazards models. We constructed a multivariable model adjusting for nine patient-level potential confounders (age, sex, household siblings, parental asthma, birth by caesarean section, breast feeding, history of systemic antibiotic use, respiratory symptoms at the time of nasal sampling and respiratory virus in age 2 month sample). These covariates were selected based on clinical plausibility and a priori knowledge.9 11 28–30 In the sensitivity analyses, we repeated the analysis by fitting a negative binomial regression model with (1) excluding infants with respiratory symptoms at the time of age 2 month nasal sampling, (2) modelling the rate of non-LRTIs (ie, ARIs not including physician-diagnosed LRTIs) during age 2–24 months as the outcome and (3) modelling the number of days with ARI symptoms during age 2–24 months as the outcome. We reported p values as two tailed, with p<0.05 considered statistically significant. Data were analysed using R version 3.5.1 with the phyloseq package for the microbiota analysis.31
Results
Study population
Of 923 infants enrolled into the ARI follow-up subcohort in the STEPS Study, 6 (0.7%) withdrew from the study. A total of 900 (97.5%) infants underwent nasal swab sampling at age 2 months (online supplementary figure E2). 16S rRNA gene sequencing of the nasal airway samples obtained 16 970 216 high-quality merged sequences, of which 16 760 908 (98.8%) were mapped to the 16S reference database. Of the 900 nasal swabs, 839 (93.2%) met the quality control requirements and had sufficient sequence depth for 16S rRNA gene sequencing (rarefaction cut-off, 1945 reads per sample) and were included in the current analyses. Among the analytic cohort, the median age at the time of nasal sampling was 2.6 (IQR 2.4–2.7) months, 52.8% were boys and 4.1% were born premature. The median age at the end of follow-up was 23.9 (IQR 17.0–24.0) months. There were no significant differences in the characteristics or clinical outcomes between infants in the analytical cohort and those without qualified microbiota data (non-analytical cohort) (online supplementary table E1).
Nasal microbiota profiles
The 16S rRNA gene sequencing of nasal microbiota at age 2 months detected 29 phyla and 911 genera. The dominant phyla were Firmicutes (52.2%), Proteobacteria (32.6%) and Actinobacteria (14.1%). The nasal airway microbiota was dominated by four genera: Moraxella (26.1%), Dolosigranulum (16.6%), Staphylococcus (16.0%) and Streptococcus (15.2%), followed by Corynebacteriaceae genus 1 (7.6%) and genus 2 (5.0%) and Neisseriaceae genus 1 (3.0%).
Clustering of nasal airway microbiota identified five distinct microbiota profiles in infants at age 2 months: (1) Moraxella-dominant (30.4%), (2) Streptococcus-dominant (22.4%), (3) Dolosigranulum-dominant (22.4%), (4) Staphylococcus-dominant (17.9%) and (5) Corynebacteriaceae-dominant (6.9%) profiles (table 1 and figure 1). The bacterial richness and alpha diversity (Shannon Index) were lowest in infants with a Moraxella-dominant profile (both p<0.001). Some of patient characteristics differed by the nasal microbiota profiles (table 2). For example, infants with a Moraxella-dominant profile were more likely to have household siblings and mild respiratory symptoms at the age 2 month visit (both p<0.05).
Richness, alpha diversity and relative abundance by nasal airway microbiota profiles at age 2 months
Characteristics and clinical outcomes in healthy children by nasal microbiota profiles at age 2 months

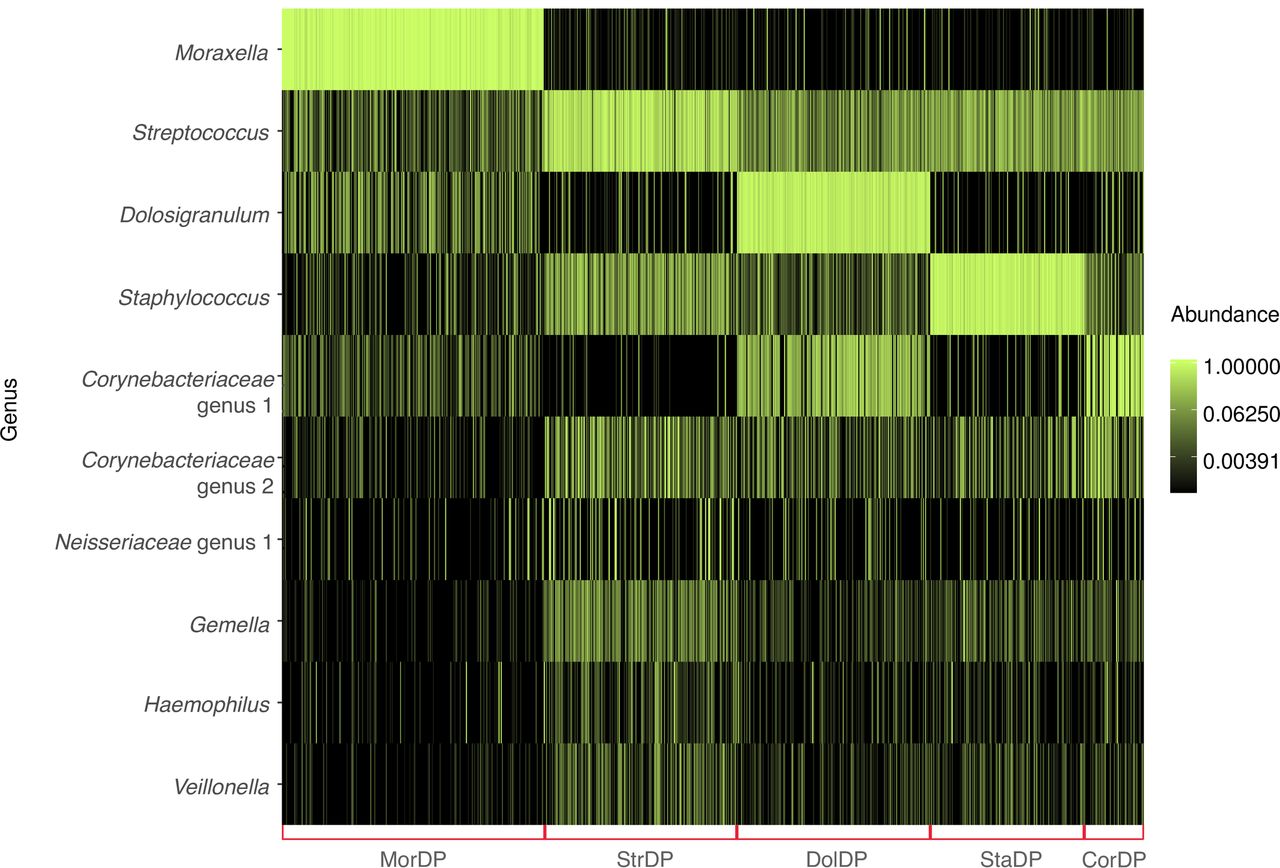{kind=link}
Clustering and composition of nasal microbiota in 839 healthy infants at age 2 months. Nasal microbiota profiles were clustered using the partitioning around medoids clustering method based on Bray-Curtis distance. The optimal number of clusters was identified by the gap statistics. Red brackets indicate five microbiota profiles: Moraxella-dominant profile (MorDP); Streptococcus-dominant profile (StrDP); Dolosigranulum-dominant profile (DolDP); Staphylococcus-dominant profile (StaDP) and Corynebacteriaceae-dominant profile (CorDP). Ten most abundant genera are shown to describe the composition of samples within the microbiota profiles.
Nasal microbiota profiles and incidence rates of ARIs
A total of 7810 episodes of ARIs (including upper ARIs and LRTIs) were documented during age 2–24 months with an incidence rate of 656 (95%CI 635 to 679) per 100 children years. During these 7810 ARI episodes, a total of 4055 physician visits and 67 hospitalisations for ARIs were documented. Children were evaluated by a physician and diagnosed with an ARI during 3233 (41%) ARIs and 59% of the ARIs were based on diary data. Altogether 285 episodes of LRTI were documented with an incidence rate of 24 (95% CI 20 to 29) per 100 children years. The duration of ARI symptoms was 4982 (95% CI 4698 to 5288) days per 100 children years. Recurrent wheezing was documented in 11.6% of children.
The incidence rate of ARIs differed between the nasal microbiota profiles. Children with an early Moraxella-dominant profile had a significantly higher incidence rate of ARIs during age 2–24 months compared with those with a Corynebacteriaceae-dominant profile (738 vs 552 per 100 children years; unadjusted incidence rate ratio (IRR), 1.34; 95% CI 1.16 to 1.54; p<0.001, table 3). After adjusting for nine potential confounders, the significant association persisted (adjusted IRR (aIRR), 1.19; 95% CI 1.04 to 1.37; p=0.01). Likewise, the incidence rate of LRTIs was significantly higher in children with an early Moraxella-dominant (aIRR, 2.79; 95% CI 1.04 to 8.09; p=0.04) and Staphylococcus-dominant (aIRR, 2.90; 95% CI 1.07 to 8.49; p=0.04) profiles, compared with those with a Corynebacteriaceae-dominant profile (table 3). In the sensitivity analyses, excluding infants with respiratory symptoms at the time of age 2 months nasal sampling (n=135), these associations persisted (online supplementary table E2). The rate of non-LRTIs (ie, ARIs not including physician-diagnosed LRTIs) was also significantly higher in children with an early Moraxella-dominant profile (online supplementary table E3). Also, the duration of days with ARI symptoms was significantly higher in children with an early Moraxella-dominant profile (aIRR, 1.61; 95% CI 1.25 to 2.06; p<0.001) compared with those with an early Corynebacteriaceae-dominant profile (online supplementary table E4). With regard to the risk of recurrent wheezing, children with an early Moraxella-dominant profile had non-significantly higher risk compared with those with an early Streptococcus-dominant profile (adjusted HR, 1.82; 95% CI 0.94 to 3.52; p=0.08; table 4 and online supplementary figure E3). Full results of the above models are shown in online supplementary tables E5-E7.
Unadjusted and multivariable-adjusted associations of nasal microbiota profiles at age 2 months with rate of acute respiratory infections (ARI) and lower respiratory infections (LRTI) during age 2–24 months*
Unadjusted and multivariable-adjusted associations of nasal microbiota profiles at age 2 months with risk of recurrent wheezing by age 24 months*
Discussion
In this prospective, population-based, birth-cohort study of 839 healthy children, we identified five nasal airway microbiota profiles at age 2 months with differential risks of subsequent ARIs. Indeed, the incidence rate of all ARIs was highest in children with an early Moraxella-dominant profile and lowest in those with an early Corynebacteriaceae-dominant profile. Similarly, incidence rates of LRTIs were highest in children with early Moraxella-dominant and Staphylococcus-dominant profiles. Our data build on previous smaller studies (n<235)9 11 reporting that certain early nasal microbiota profiles are associated with increased susceptibility to ARIs (‘risk microbiota’), and extend them by demonstrating robust longitudinal microbiota-ARI associations in a large well-characterised cohort study.
Recent studies have suggested that early Moraxella-dominant airway microbiota may be associated with increased risk of ARIs but the findings have not been consistent. In a cohort of 234 Australian infants with a high risk of atopy, Teo and colleagues reported that early Moraxella-dominant nasopharyngeal microbiota was associated with an earlier first upper respiratory infection.11 Similarly, in a cohort of 112 children in the Netherlands, Bosch and colleagues reported an association between enrichment of Moraxella in early life in children and a high frequency of ARIs during the first year of life.9 In contrast, in an analysis of trial data in the Netherlands (n=60), Biesbroek and colleagues found that infants with Moraxella-dominant nasopharyngeal microbiota had a lower risk of subsequent ARI.7 However, the study by Biesbroek and colleagues used retrospective parent-reported ARI as a dichotomised outcome, and the rate of ARIs—a frequent outcome in infants and young children—was not assessed. These differences in methodology may explain the inconsistent results. Earlier studies have also linked detection and dominance of airway microbiota by Moraxella at the time of ARI to the symptoms11 13 and severity11 of ARIs in children. However, we show that the longitudinal association between early Moraxella-dominant airway microbiota and increased risk of ARIs and LRTIs persisted when excluding infants with respiratory symptoms at the time of nasal sampling. Based on our results and previous studies, colonisation with Moraxella in the airway is likely to play an important role in the susceptibility to ARIs in young children.
We also demonstrated that children with an early Moraxella-dominant nasal microbiota had the highest rate of LRTIs. Previous data on early airway microbiota assessed by next-generation sequencing methods and rates of LRTIs are scarce. The Australian study found that Moraxella enrichment at the time of ARI was significantly associated with higher risks of LRTI.11 In contrast, in case-control studies, Moraxella-dominant airway microbiota has been found to be associated with lower risks of severe bronchiolitis8 and RSV hospitalisation,12 and not to be associated with pneumonia in children.13 However, in these studies, the nasal microbiota was investigated at the time of severe infection (with possible dysbiosis), which may explain the difference in results. Our finding that early Moraxella-dominant airway microbiota may play a role in the risk of developing LRTIs is supported by previous reports using bacterial culture or PCR. By using culture, in the Copenhagen Prospective Study on Asthma in Childhood (COPSAC) cohort of 321 Danish children born to mothers with asthma, early colonisation at age 1 month with Moraxella catarrhalis was associated with first wheeze and hospitalisation for wheeze.32 In a later subanalysis of the same study (n=265), early airway colonisation with M. catarrhalis was associated with increased risks of pneumonia and bronchiolitis in the first 3 years of life.14 Similarly, in a Danish study using PCR (n=218), neonatal nasal colonisation with Streptococcus pneumoniae, Haemophilus influenzae or M. catarrhalis was associated with cough and wheezing episodes in the first year of life.33 Potential reasons for discrepancies across studies include differences in patient populations, settings (eg, hospital vs community setting), sampling methods (eg, nasal swab vs nasopharyngeal aspirate) and laboratory techniques (eg, culture vs next-generation sequencing). The validity of our results is buttressed by the use of 16S rRNA gene sequencing of the nasal microbiota from the population-based cohort of healthy children and a sample size that is much larger than any prior study on this topic.
There are several potential mechanisms that explain the observed association of early nasal airway microbiota with susceptibility to ARIs and LRTIs. It is possible that there is a causal relationship—for example, Moraxella alters the mucosal immune responses and increases susceptibility to ARIs and LRTIs. Indeed, M. catarrhalis colonisation of the airways in asymptomatic neonates has been shown to induce species-specific stimulation of inflammatory mediators (eg, a mixed T helper cell (TH) type 1/TH2/TH17 response with high levels of interleukin 1b, tumour necrosis factor-α and macrophage inflammatory protein-1β).5 One possibility is that early Moraxella-dominant nasal microbiota may be only a marker of children with increased susceptibility to ARIs or LRTIs. Furthermore, a recent study reported that Moraxella interacts with respiratory viruses (eg, rhinovirus), contributing to the symptoms and severity.34 Children with an early Moraxella-dominant nasal microbiota profile were more likely to have household siblings. It is possible that this pattern of nasal microbiota was transferred from the sibling. Nevertheless, in the multivariable models, the early Moraxella-dominant nasal microbiota profile was associated with increased rates of ARIs and LRTIs independent from sibling status. Additionally, as the early Corynebacteriaceae-dominant profile was associated with the lowest rates of ARIs and LRTIs, it is possible that Corynebacteriaceae confer resistance against respiratory pathogens and thus against ARIs (‘resilience microbiota’). Previous reports have also linked airway Corynebacteria to lower risks of ARIs and wheeze in children.7–9 13 35 Ultimately, we may learn that these different mechanisms are not mutually exclusive. Our data, in conjunction with the previous studies, suggest a complex interplay between airway microbiota, respiratory viruses and host immune response in the development and morbidity of ARIs.12 36
Our study has potential limitations. First, while ARI may involve the lower airways, our data relied on the nasal airway microbiota. However, the nasal sampling is a non-invasive procedure in young infants. Previous studies have also reported a correlation between upper and lower airway microbiota.37–39 Second, as we used nasal samples of healthy infants with potentially low amounts of bacterial DNA, part of the nasal samples had insufficient sequence depth and were excluded from the analyses. Nonetheless, nasal samples were analysed from >90% of the infants and the analytical cohort did not differ from the infants without microbiota data. Third, the study design precluded us from examining succession of the airway microbiota, and its association with respiratory health in children. To address this question, the cohort is currently being followed longitudinally with multiple specimen collections; we would like to test for nasal microbiota at different time points in childhood. Fourth, the definition of ARI was based on diary in 59% of ARIs. Symptoms of ARI may be subjective with some parents being more cautious in reporting milder symptoms than others. However, the documented rate of ARIs in this study is consistent with the literature.2 3 Furthermore, we also examined physician-diagnosed LRTIs, and found a larger effect size in the association between the early Moraxella-dominant microbiota profile and the LRTI outcome, which supports the validity of our inferences. Fifth, despite the statistical adjustment in the multivariable models, the observed associations do not prove causality and may be explained by unmeasured confounders, such as environmental microbiota. Sixth, the number of patients with recurrent wheezing in this healthy child cohort was relatively low, leading to a limited statistical power to examine the association with recurrent wheezing. Lastly, with 16S rRNA gene sequencing methods, identification at the species level was not possible in the present study. However, by applying metagenomic whole-genome shotgun sequencing to nasopharyngeal samples of infants hospitalised for bronchiolitis, we previously demonstrated that Moraxella genus was dominated by M. catarrhalis in infant airways.36
Conclusions
In this prospective, population-based birth cohort study, we identified five distinct nasal microbiota profiles in healthy 2-month-old infants. Early Moraxella-dominant nasal microbiota was associated with a significantly increased rate of all ARIs as compared with those with an early Corynebacteriaceae-dominant profile. Rates of LRTIs were significantly higher in children with early Moraxella-dominant and Staphylococcus-dominant profiles. These findings suggest that early nasal microbiota may play an important role in the development and severity of ARIs in children. Our data should advance research into understanding the complex interplay among the immunity, airway microbiome and pathobiology of ARI in young children, and characterising the children at higher risk for ARIs.
Acknowledgments
The authors thank all the families who participated in this study, the midwives for their help in recruiting the families and the whole STEPS Study team for assistance with data collection, study nurses Niina Lukkarla, Petra Rajala and Mira Katajamäki for their assistance in the study clinic, Tiina Ylinen for her technical assistance in virus detection, and Anne Kaljonen for her assistance with data handling. They thank Finnish Functional Genomics Centre, supported by University of Turku, Åbo Akademi University, and Biocenter Finland, for DNA extractions, and Kristi Hoffman for her assistance with the 16S rRNA gene sequencing analysis.
References
Footnotes
Contributors LT collected the data, obtained the funding, carried out the statistical analysis, drafted the initial manuscript and approved the final manuscript as submitted. KH carried out the statistical analysis, reviewed and revised the initial manuscript, and approved the final manuscript as submitted. MW carried out the virological analysis and DNA extractions, reviewed and revised the manuscript, and approved the final manuscript as submitted. NA and JFP generated the microbiome data, carried out the initial statistical analysis, reviewed and revised the manuscript, and approved the final manuscript as submitted. CAC critically reviewed and revised the initial manuscript, and approved the final manuscript as submitted. VP conceptualised and designed the study, obtained the funding, supervised the conduct of the study, reviewed and revised the manuscript, and approved the final manuscript as submitted.
Funding This work was supported by the University of Turku; the Abo Akademi University; the Turku University Hospital; the Academy of Finland (Grant nos. 123571, 140251 and 277535); the Finnish Medical Foundation; the Päivikki and Sakari Sohlberg Foundation; the Foundation for Pediatric Research; Research Funds from Specified Government Transfers, Hospital District of Southwest Finland; the Foundation of the Finnish Anti-Tuberculosis Association; the Tampere Tuberculosis Foundation; the Allergy Research Foundation; the Emil Aaltonen foundation; the Maud Kuistila Memorial Foundation; the Orion Research Foundation; the Paulo Foundation; the Väinö and Laina Kivi Foundation and the Finnish Cultural Foundation.
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The STEPS study was found ethically acceptable by the Ministry of Social Affairs and Health (STM 1575/2008, STM 1838/2009) and the Ethics Committee of the Hospital District of Southwest Finland (19.2.2008 §63, 15.4.2008 §134, 19.4.2011 §113).
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Data are available upon reasonable request.